 |
Kaskelotens väldiga huvud innehåller en hjärna som kan vara den största som någonsin funnits hos något djur. Men den största delen av huvudet upptages av det märkliga spermacetiorganet. Detta organ väger flera ton och utgör cirka 90 procent av huvudets vikt. Courtesy of the IFAW Charitable Trust. |
|
Kaskeloten (även kallad spermacetivalen) har ett oproportionerligt stort huvud. Det kan utgöra en fjärdedel av valens längd och en tredjedel av dess vikt. Om man tittar på valens skelett ser man en relativt liten skalle i huvudets bakre del och käkar i form av en lång smal näbb längs huvudets undersida. Men huvudet på en levande val är stort och nästan fyrkantigt. Vad är det som döljer sig i kaskelotens huvud?
Kaskeloten hör till tandvalarna, tillsammans med bland annat späckhuggare och delfiner. Den finns i alla stora oceaner. Hannarna kan bli 18 meter långa och väga 50 ton. Honorna är mindre. Kaskeloten är ett rovdjur som huvudsakligen lever på bläckfiskar. Det är inte ovanligt att en kaskelot, som dyker efter föda, stannar under vattnet i 50 minuter och då dyker ner till 1 000 meters djup. F–rmodligen kan kaskeloter göra dyk som varar upp till cirka 2 timmar och når ner till 2 000-3 000 meters djup. Läs om dykrekord hos däggdjuren på en annan sida.
I huvudet finns ett stort organ fyllt med en sorts olja. Oljan kallas spermacetiolja och organet följaktligen spermacetiorganet. Se bilden nedan. Organet består av två skilda strukturer. Överst finns en oljefylld spermacetisäck ("case") omgiven av ett tjockt och starkt fodral av bindvävnad. Under denna finns flera av seg bindvävnad avgränsade fack fyllda av spermacetiolja. Denna del av organet kallas "junk".
Spermacetioljan användes förr bland annat som smörjmedel och som bas i kosmetiska krämer. Den ska inte förväxlas med ambran som bildas i kaskelotens tarmar och används i parfymer.
En nyckel till spermacetiorganets funktion är att förstå vad valen använder sina asymmetriska näsgångar till. Längst fram och längst upp på huvudet har valen sitt blåshål. Se bilden nedan. Från blåshålet utgår två näsgångar. Den vänstra går vid sidan av spermacetiorganet bakåt och neråt genom huvudet. Det är genom denna gång som valen andas. Den högra näsgången går genom den nedre delen av spermacetisäcken. Den är cirka 1 meter bred, men tillplattad i höjdled och därför mycket lägre. Den kan vara cirka 5 meter lång. I både framänden och bakänden vidgas den och bildar luftfyllda säckar vilka täcker de främre och bakre ändarna av spermacetiorganet. De båda näsgångarna är förenade med varandra via en smal gång nära deras bakre ändar. Baktill, nära den bakre luftsäcken, finns det en förbindelse mellan spermacetisäck och "junk"-en (ej utritad på figuren nedan).
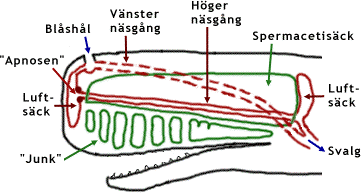 |
Kaskelotens huvud i genomskärning. Bilden visar schematiskt ett lodrätt längdsnitt mitt i valkroppen. Den rödstreckade vänstra näsgången löper på huvudets vänstra sida bredvid spermacetiorganet och befinner sig således framför bildens plan. Bilden förklaras vidare i texten. |
|
I den främre säcken finns en öppning som leder till fortsättningen av den högra näsgången. Där sitter en struktur som kallas "museau de singe", eller "apnosen". Den ser som två hårt sammanpressade läppar, lite som orangutangens mun. Man tror att ljud produceras genom att luft pressas med högt tryck fram genom högra näsgången till apnosen som då bringas i vibration av luftströmmen. Detta skulle ge upphov till kaskelotens karakteristiska klickljud. Dessa ljud använder kaskeloten vid dykning, sannolikt för att orientera sig med hjälp av sonar (ekolokalisering), läs om hur delfiners sonar fungerar på en annan sida. Kaskeloten lokaliserar föremål med ledning av de ekon som klickljuden ger upphov till när de träffar något föremål. Valen sänder ut en serie klickljud och av de ekon som kommer tillbaka får den en "ljudbild" av sin omgivning. Ekoljuden leds sannolikt genom en fettfylld hålighet i underkäken till innerörat, inte genom en hörselgång som hos de flesta andra däggdjur.
Kaskeloten behöver verkligen en sonar för det är totalt mörkt på det djup där den jagar. Kaskeloterna producerar också andra ljud än klickljuden. En del av ljuden har förmodligen sociala funktioner som signaler kaskeloter emellan. Nederst på denna sida kan du lyssna på kaskelotljud.
Det finns flera hypoteser om hur ljudet rör sig genom spermacetiorganet. Det är nämligen svårt att studera kaskeloter. Man är hänvisad till att studera hur riktat ljud från högtalare leds genom nosen hos strandade döda kaskeloter. Enligt den sannolikaste hypotesen leds sonarljudet från apnosen först bakåt genom spermacetisäcken, alltså inte framåt genom den främre luftsäcken och ut i vattnet. Det ljud som leds framåt reflekteras troligen i stället mot den främre luftsäcken. Ljudet som leds bakåt genom spermacetisäcken reflekteras mot den luftfyllda bakre luftsäcken och leds genom "junk"-en framåt och ut i vattnet. De oljefyllda facken i "junk"-en skulle då fungera som akustiska linser och ljudet skulle kunna ledas framåt genom "junk"-en, utan att det läcker ut från den i sidled. "Junk"-en skulle alltså kunna fokusera ljudet så att en koncentrerad stråle av ljud leds genom dess främre ände ut i vattnet. Hela spermacetiorganet skulle således fungera som ett böjt blåsinstrument, till exempel en fagott eller en tenorsaxofon. Kanske är spermacetiorganet den effektivaste sonarapparaten hos tandvalarna. Enligt teoretiska beräkningar skulle en kaskelot med sin sonar kunna identifiera en bläckfisk på flera hundra meters avstånd.
Ljuden som skapas i apnosen ger sannolikt även upphov till ekon inne i spermacetisäcken. Ljudet som reflekteras från den bakre luftsäcken kan inte bara ledas framåt genom "junk"-en. De kan också ledas framåt genom spermacetisäcken och reflekteras mot den främre luftsäcken för att återigen reflekteras mot den bakre luftsäcken eller lämna valen genom "junk"-en. På så sätt skulle ljudet kunna studsa fram och tillbaka fler gånger i spermacetisäcken mellan de båda luftsäckarna. Luftsäckarna fungerar som reflektorer just på grund av att de innehåller luft. Det vanligaste kaskelotljudet är en serie av klick som avges med oförändrade tidsmellanrum och successivt avtar i ljudstyrka. Detta mönster överensstämmer mycket väl med ekolokaliseringshypotesen. Tidsmellanrummen mellan klicken skulle då motsvara den tid det tar för ljudet att färdas en gång fram och tillbaka genom spermacetisäcken.
Det har spekulerats att spermacetisäckens främsta funktion är att framställa sonarklick med konstanta tidsmellanrum. Detta skulle kunna ge valen en mer exakt uppfattning om hur omgivningen ser ut och göra det lättare för den att lokalisera sitt byte. En annan funktion skulle kunna vara att en kaskelot kan ge andra valar information om hur stor den är. En stor val har större avstånd från den främre luftsäcken till den bakre och därför ett något längre tidsavstånd mellan klicken. Detta skulle kunna vara av betydelse när hanar konkurrerar om en hona.
Kaskeloten kan faktiskt producera ljud med högre ljudstyrkor än man uppmätt från något annat djur, läs om kaskelotljudens enorma ljudstyrka på en annan sida.
 |
Kaskelotens utandning, den så kallade blåsten. Blåsten kan se olika ut hos olika valarter och ger viss hjälp vid artbestämningen ute på havet. Valarna andas ut och in genom det så kallade blåshålet som sitter på huvudets ovansida. Hos tandvalar utgörs blåshålet av en enda öppning, medan bardvalarna har behållit två öppningar svarande mot våra näsborrar. Courtesy of the IFAW Charitable Trust. |
|
Det torde stå klart att den viktigaste funktionen med spermacetiorganet är att producera och rikta ljud. Det står också klart att organet är en modifierad version av andra tandvalars sonarapparat. Men det finns andra hypoteser när det gäller kaskeloten. Vissa av dessa hypoteser skulle kunna återspegla det faktum att spermacetiorganet även har andra sekundära funktioner, andra hypoteser är mindre sannolika.
Ett fantasifullt förslag är att kaskeloten skulle producera ett så starkt ljud att bytesbläckfiskarna blir medvetslösa av den enorma ljudstyrkan och därmed ett lätt byte för valen. Detta skulle kunna förklara kaskelotens förmåga att fånga mycket stora bläckfiskar, trots att den har en så liten underkäke. Stödet för denna så kallade "big bang"-hypotes är svagt. Men den kan inte uteslutas.
En annan hypotes är att valen aktivt värmer eller kyler oljan i sitt spermacetiorgan för att ge sig själv ökad eller minskad flytförmåga. När valen vilar vid ytan håller spermacetioljan en temperatur på 33 °C. Vid tester på oljan har man funnit att oljan börjar stelna när dess temperatur sjunker under cirka 30 °C. Detta minskar oljans volym, det vill säga ökar dess densitet ("täthet"). Valen skulle, om den kyler sin olja, få högre densitet än vattnet, som omger den, och därmed skulle den sjunka. Det blir då lättare för den att dyka ner till sina jaktmarker som ligger på åtskilliga hundra meters djup. Kylningen skulle kunna ske genom att valen minskar blodflödet i spermacetiorganet och låter det omgivande vattnet kyla ner oljan i organet. Värme skulle kunna transporteras till huvudets yta av blodet och där avges till omgivningen. Valen skulle också kunna ta in kallt vatten genom den breda högra näsgången och på så vis kyla spermacetiorganet inifrån. Om valen har dykt djupt ner och vill stiga upp, skulle den kunna värma den stelnade oljan genom att öka blodflödet till spermacetiorganet. Detta skulle ge en lägre densitet hos oljan och ökad flytförmåga.
Flera iakttagelser talar emellertid emot att spermacetiorganet reglerar flytförmågan. Det är alltför glest mellan blodkärlen i spermacetiorganet för att värme effektivt ska kunna transporteras med blodet ut ur och in i organet. Man skulle också förvänta sig att det fanns parallella utåtgående och inåtgående blodkärl som skulle fungera som värmeväxlare. Inga sådana strukturer har iakttagits. Utan värmeväxlare skulle det bli alltför svårt att kyla spermacetiorganet. Djur som håller vissa kroppsdelar kallare än andra brukar använda sig av värmeväxlare, läs om hur dessa värmeväxlare fungerar på en annan sida.
Kaskelothanar är större än honorna och har också ett större spermacetiorgan, både i absoluta mått mätt och i förhållande till kroppsvikten. Detta kan mycket väl vara ett exempel på så kallad sexuell selektion. En hane med stort spermacetiorgan skulle kunna göra honorna imponerade och mera parningsvilliga. Dessutom har man iakttagit att hanar kämpar med varandra genom att sätta fart och simma på varandra med nosen. De stångas alltså precis som partåiga hovdjur. Man kan föreställa sig vilka enorma krafter det handlar om. Märkligt nog är de partåiga hovdjuren, jämte flodhästarna, valarnas närmaste släktingar bland däggdjuren. Spermacetiorganet skulle alltså ha samma funktion som hornen bland hovdjuren. Hypotesen stöds bland annat av att huden är mycket tjock framtill på kaskeloternas nos och av att gamla kaskelothanar ofta har ärr på huvudet.
Kaskelothanarnas benägenhet att stångas skulle också förklara flera dokumenterade fall, då kaskeloter rammat fartyg med nosen. Två skepp blev så skadade att de sjönk, Essex (1821) och Ann Alexander (1851). Dessa händelser inspirerade författaren Herman Melville när han skrev sin klassiska roman "Moby-Dick" (1851). I romanen är den tyranniske kapten Ahab på sin båt Peqoud ute efter att hämnas på den vita kaskeloten Moby-Dick, som lemlästat honom under en tidigare valfångstfärd. Till slut får kapten Ahab harpunlinan om halsen när han ska harpunera valen och dras ner i djupet av Moby-Dick. Moby-Dick rammar sedan Peqoud så att båten börjar sjunka. Endast bokens berättare, som kallar sig Ishmael, överlever skeppsbrottet.
| Klick |
Klingande |
Trumpetljud |
| Några av kaskeloternas ljud. De vanliga klicken anses vara ekolokaliseringsljud. De klingande ljuden tycks produceras av vuxna hannar och kan vara sociala signaler. Trumpetljuden uppkommer kanske när valen rensar näsgångarna (d.v.s. nyser). Men man vet inte mycket om ljudens funktioner. Courtesy of the IFAW Charitable Trust (copyright Steve Knight). |
Referenser
På grund av nya rön har texten uppdaterats och utökats år 2013 av Anders Lundquist.
D.R. Carrier, S.M. Deban och J. Otterstrom: The face that sank the Essex: potential function of the spermaceti organ in aggression (Journal of Experimental Biology 205:1755‚1763, 2002).
M.R. Clarke: Stucture and proportions of the spermaceti organ in the sperm
whale (J. Marine Biol. Assoc. U.K. 58:1-17, 1978).
M.R. Clarke: Buoyancy control as a function of the spermaceti organ in the
sperm whale (J. Marine Biol. Assoc. U.K. 58:27-71, 1978).
M.R. Clarke: The head of the sperm whale (Scientific American 240:106-117, 1979).
J.C. Goold, J.D. Bennell och S.E. Jones: Sound velocity measurements in the
spermaceti oil under the combined influences of temperature and pressure (Deep-Sea
Research I 43:961-969, 1996).
T.W. Cranford: The spem whale's nose: sexual selection on a grand scale? (Mammal Marine Science 15:1133-1157, 1999).
J.C. Goold och S.E. Jones: Time and frequency domain characteristics of sperm
whale clicks (Journal of the Acoustical Society of America 98:1279-1291, 1995).
IFAW Charitable Trust.
R.S. Mackay: A theory of the spermaceti organ in sperm whale sound production (Ur: R.G. Bunsel och J.F. Fish (red) Animal Sonar Systems. Plenum, New York, 1980, sid 937-940.).
P.T. Madsen, D.A. Carder, W.W.L. Au, P.E. Nachtigall, B. Møhl och S.H. Ridgway: Sound production in neonate sperm whales (Journal of the Acoustical Society of America 113:2988-2991, 2003).
P.T. Madsen, R. Payne, N.U. Kristiansen, M. Wahlberg, I. Kerr och B. Møhl: Sperm whale sound production studied with ultrasound time/depth-recording
tags(Journal of Experimental Biology 205:1899‚1906, 2002).
B. Møhl, P.T. Madsen och M. Wahlberg: Sound transmission in the spermaceti complex
of a recently expired sperm whale calf (Acoustics Research Letters Online DOI 10.1121/1.1538390, 2002).
Till början på sidan
Till "Artiklar om djur"
|


