 |
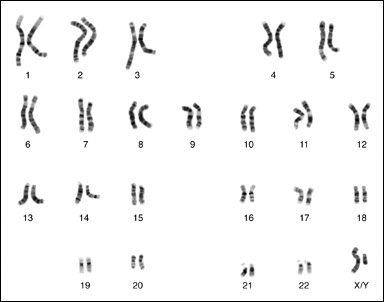
Människans 46 kromosomer, ordnade parvis. I könscellerna, det vill säga äggcellerna och spermierna, finns det ett exemplar av varje kromosom. I alla andra celler finns två exemplar av varje kromosom, med ett undantag. Män har två olika könskromosomer, X och Y. Kvinnor har däremot två likadana könskromosomer, båda X. Deras ena X-kromosom inaktiveras dock mellan celldelningarna till en så kallad "Barr body". Därmed får generna i X-kromosomen lika stor effekt hos båda könen. Bildens kromosomer är från en man. Längst ner till höger ser man Y-kromosomen tillsammans med den större X-kromosomen. Courtesy of the National Human Genome Research Institute (USA), in the public domain. |
|
Vad är det som avgör om ett foster blir hona eller hane, flicka eller pojke. Förvånande nog är skillnaderna mycket stora mellan olika djurgrupper. Hos vissa ryggradsdjur, bland dem människan, bestäms könet av genetiska skillnader mellan honfoster och hanfoster. Hos andra bestäms könet märkligt nog av miljön, vanligen av den temperatur som fostret utsätts för.
Könet är inte bestämt i tidiga embryon

Hos tidiga embryon av ryggradsdjur är könet ännu inte bestämt. Så småningom bildas ett par genitalkammar i kroppshålans bakre vägg. Dessa kammar innehåller en bark och en märg. Barken kan ge upphov till äggstockar (ovarier). Märgen kan ge upphov till testiklar. Föregångarna till äggcellerna och spermierna härstammar märkligt nog inte från fostret, utan från gulesäcken vägg, alltså det cellskikt som omger äggets gula. Därifrån vandrar dessa så kallade primordiella könsceller till genitalkammarna. Alla rygggradsdjur har en gulesäck, förvånande nog även däggdjuren. Däggdjurens gulesäck innehåller emellertid ingen gula, utom hos de äggläggande kloakdjuren. Läs om fosterhinnor på en annan sida.
Könsbestämning med könskromosomer
Könet kan bestämmas genetiskt, ofta av särskilda könskromosomer i arvsmassan. Hos människan och nästan alla andra däggdjur har hanar en X-kromosom och en Y-kromosom, honor två X-kromosomer. Hos honor inaktiveras dock den ena X-kromosomen, vilket gör att båda könen producerar lika mycket av denna kromosoms genprodukter. Den inaktiverade X-kromosomen, könskromatinet ("Barr body") kan iakttagas i honliga cellkärnor. Hos fåglar har däremot hanar två Z-kromosomer, honor en Z-kromosom och en W-kromosom. Texten fortsätter under bilden.
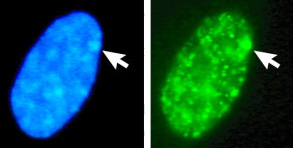 |
Pilarna visar könskromatinet ("Barr body") från en kvinna i två cellkärnor, som färgats på olika sätt. Könskromatinet är en inaktiverad X-kromosom. Inaktiveringen av kvinnors ena X-kromosom gör att både kvinnor och män bara har en aktiv X-kromosom i sina celler. Därmed blir effekterna av denna kromosoms gener lika stora hos båda könen, så kallad doskompensation. Doskompensation förekommer hos däggdjur, men inte hos fåglar. Intressant nog är det sannolikt just frånvaron av doskompensation som bestämmer könet hos fåglar, mera därom längre ner på denna sida. Courtesy of S. M. Gartler et al. under this CC License. |
|
Temperaturbetingad könsbestämning
Könet kan också bestämmas av miljöfaktorer, vanligen av den temperatur som det utvecklande embryot utsätts för under en kritisk period. Oftast finns det då en tröskeltemperatur. Vid temperaturer ovanför tröskeln utvecklas embryot till det ena könet, vid temperaturer under tröskeln till det andra. Hos vissa arter uppkommer hanar vid höga och honor vid låga temperaturer, hos andra hanar vid låga och honor vid höga. Åter andra arter har två tröskeltemperaturer. Då uppkommer hanar vid temperaturer mellan tröskelvärdena, honor både vid högre och vid lägre temperaturer.
Temperaturbetingad könsbestämning förekommer hos alla krokodildjur, hos de flesta sköldpaddor, hos några ödlor och ormar samt hos många fiskar. Kräldjur lägger ofta äggen i nästen. Krokodiler täcker över dem med växtdelar, havssköldpaddor gräver ner dem i sanden på havsstränder. Det temperaturspann, inom vilket både hanar och honor kan kläckas i ett näste, är mycket smalt. Om båda könen kläcks finns det förmodligen små temperaturvariationer inom nästet. Hos havssköldpaddor har man visat att vissa nästen bara ger upphov till honor, andra till hanar. Texten fortsätter under bilden.
 |
Ägg av en havssköldpadda. Honan har grävt ett hål i strandens sand och lagt äggen, men ännu inte fyllt igen hålet. Hos de flesta sköldpaddor bestäms könet av den temperatur embryona i äggen utsätts för. Detta innebär att endast hanar kan kläckas ur vissa nästen, endast honor ur andra. Courtesy of Steve Hillebrand and the U.S. Fish and Wildlife Service, in the public domain. |
|
Andelen hanar och honor hos olika djur
Vid genetisk könsbestämning ligger könskvoten i regel nära 0,5, det vill säga 50 procent. Detta innebär att andelen hanar är ungefär 50 procent och andelen honor därmed ungefär lika stor. Hos människan föds cirka 52,5 procent pojkar och cirka 47,5 procent flickor. Orsaken till detta är oklar, man kan bara spekulera. En möjlig orsak är att spermier med Y-kromosomer simmar något lite snabbare än de med X-kromosomer och därmed har större chans att nå ägget först. Å andra sidan är dödligheten något större hos det manliga könet vid alla åldrar efter födelsen och mäns medellivslängd kortare än kvinnors.
Det är synnerligen märkligt att könet kan bestämmas av temperaturen. Vid temperaturbetingad könsbestämning kan nämligen könskvoten variera mycket mer än vid genetisk. Man kan tycka att det naturliga urvalet borde leda till 50 % honor och 50 % hanar. Den varierande könskvoten vid temperaturbetingad könsbestämning kan naturligtvis orsakas av väderlekens temperaturvariationer. Teoretiskt skulle detta dock kunna vara både en fördel och en nackdel. Men om detta vet man mycket lite. Om populationen är liten skulle det kunna tänkas vara en fördel med en större andel honor. Då skulle populationen kunna växa snabbare. Det skulle vara mycket intressant att veta om krokodilhonor och sköldpaddshonor kan välja nästets läge efter den temperatur som råder där.
Man har befarat att den globala uppvärmningen skulle ställa till problem för djur med temperaturbetingad könsbestämning. Också här är kunskapsläget oklart. Å ena sidan har dessa djur upplevt stora klimatiska temperaturvariationer under sin evolution. Å andra sidan värms jordklotet nu upp snabbare än någonsin under livets utveckling. I en artikel från 2018 har man påvisat ett extremt överskott av honor från vissa äggläggningsstränder för soppsköldpaddan. Man tror att detta beror på den globala uppvärmningen och befarar att endast honor kommer att kläckas på dessa stränder i framtiden.
Hur fungerar könsbestämningen hos människan och andra däggdjur?
Hos människan och andra äkta däggdjur (placentadäggdjur) finns det i de blivande könsorganen två komplexa mekanismer, varav den en leder till honlig och den andra till hanlig utveckling. Det handlar om genetiska kedjereaktioner, i vilka proteinprodukter av gener aktiverar andra gener. Mycket grovt förenklat skulle man kunna säga att utveckling till hona är grundtillståndet och att Y-kromosomen stimulerar hanlig utveckling, samtidigt som den hämmar honlig utveckling. Texten fortsätter under bilden.
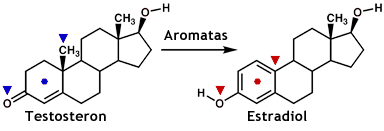 |
Små skillnader kan ge stora effekter. Bilden visar de kemiska formlerna för två könshormoner hos människan. Testosteron är det viktigaste av de androgener som insöndras från testiklarna. Estradiol är det viktigaste av de östrogener som insöndras från äggstockarna. Skillnaderna mellan de två molekylerna är markerade med pilspetsar och prickar. Intressant nog är bildning av könshormoner inte det första steget, när könet bestäms hos ett människofoster. Det viktiga enzymet aromatas katalyserar omvandlingen av testosteron till estradiol.
För de kemiskt bevandrade följer här en detaljerad beskrivning. De kolatomer som finns i sexhörningarnas och femhörningarnas hörn är inte utritade. De väteatomer som binder till kolatomerna är inte heller utritade. Testosteron bär en ketogrupp där estradiol bär en hydroxylgrupp. En metylgrupp saknas hos estradiol. Dessa båda förändringar leder till att den vänstra sexringen aromatiseras och får förändrade egenskaper. From Wikimedia Commons, in the public domain. |
|
På Y-kromosomen finns en överordnad gen som kallas Sry. Dess proteinprodukt binds till ett annat protein i cellkärnan hos de celler som senare ger upphov till testiklarnas så kallade Sertoliceller. Det bildade proteinkomplexet aktiverar en annan gen, Sox9. Via komplexa mekanismer leder detta till hanlig utveckling och hämning av honlig utveckling. Hos honor saknas Y-kromosomen och därmed genen Sry. Hos dem hämmas genen Sox9, i stället för att aktiveras. Detta sker med hjälp av proteinprodukten från genen Fox12 i de celler som senare ger upphov till äggstockarnas så kallade follikelceller. Via komplexa mekanismer leder detta till honlig utveckling.
Innan könet bestämts bildas i fostret två gångsystem, de Wolffska och de Müllerska gångarna. Hos hanfoster utvecklas de Wolffska gångarna till sädesledarna, sädesblåsorna och bitestiklarna, medan de Müllerska degenererar och bara finns kvar som rudiment. Hos honfoster utvecklas de Müllerska gångarna till äggledarna, livmodern och slidan, medan de Wolffska gångarna degenererar och bara finns kvar som rudiment. Läs om rudiment på en annan sida.
Hos hanliga foster insöndras så småningom manliga könshomoner, androgener, framför allt testosteron, från testiklarna. Notera emellertid att det inte är könshormoner, androgener hos män och östrogener hos kvinnor, som är det första steget i de kedjerektioner som bestämmer könet. Det är närvaron eller frånvaron av Y-kromosomens Sry-gen, och därmed av Sry-proteinet, som är avgörande. Androgener är emellertid nödvändiga för de inre och yttre könsorganens fortsatta utveckling i hanliga foster. Östrogener är däremot inte nödvändiga för könsorganens utveckling i honliga foster.
Effekter av androgener och östrogener efter födelsen hos människor
Efter födelsen är androgener och östrogener, jämte andra hormoner, nödvändiga för pubertetsutvecklingen, menstruationscykeln och spermieproduktionen. Dessutom ger de upphov till de så kallade sekundära könskaraktärerna, till exempel skillnaderna i kroppsform och kroppsbehåring mellan kvinnor och män. Slutligen påverkar de hjärnan, vilket leder till skillnaderna i könsdrift och sexuellt beteende mellan män och kvinnor. Märkligt nog, påverkas könsdriften hos kvinnor troligen mest av androgener från binjurarna.
Hos de flesta däggdjur orsakar androgener helt klart aggressivt beteende hos hanar, särskilt när de konkurrerar om honor i samband med parningen. Även hos människan påverkar androgener och östrogener mycket sannolikt känslor och beteenden, dock på ett komplicerat sätt och i samverkan med andra genetiska faktorer och med miljöfaktorer. Många sådana beteenden är dock svåra att studera hos människan, bland annat beroende på varierande hormonhalter under loppet av kvinnors menstruationscykel och varierande hormonhalter i olika situationer. Dessutom finns det individuella skillnader. Mäns jämfört med kvinnors större benägenhet framför allt till dominansbeteende, men därmed också till aggression och våld, torde dock till stor del orsakas av androgener, även om andra faktorer bidrar. Effekten av testosteron hos män verkar till exempel vara olika hos olika personlighetstyper.
Könsbestämning hos fåglar
Hos nästa alla fåglar bestäms könet av könskromosomer. Som ovan nämnts, har hanar två Z-kromosomer, honor en Z-kromosom och en W-kromosom. Könet bestäms under fosterutvecklingen. Hur detta sker är oklart. Enligt en hypotes bestäms könet av genen DMRT1 som finns på Z-kromosomer. Denna gen kodar för ett protein, som är en transkriptionsfaktor. Transkriptionsfaktorer aktiverar eller inhiberar andra gener. Eftersom hanembryon har två DMRT1-gener får de en högre halt av DMRT1-faktorn än honor i sina så kallade odifferentierade gonader. Kanske är det denna skillnad som avgör om en odifferentierad gonad utvecklas i hanlig riktning till en testikel eller i honlig riktning till en äggstock. Slår man ut DMRT1-genen hos hanembryon får de äggstocksliknande gonader, om än inte normala äggstockar. Dessutom aktiveras enzymet aromatas, som omvandlar det hanliga könshormonet testosteron till det honliga könshormonet estradiol. Texten fortsätter under bilden.
 |
Den spelande tjädertuppens utmanande uppträdande är betingat av manliga könshormon, androgener. Det finns en stor mängd studier som påvisat effekter av könshormoner på däggdjurs och fåglars beteende. Courtesy of A. Neumann, from Encyclopedia of Life under Creative Commons Attribution-NonCommercial 3.0 Unported License. |
|
Könskromosomerna styr det grundläggande och första steget vid könsbestämningen. Deras aktivitet är oberoende av hanliga och honliga hormoner. Deras aktivitet leder emellertid till aktivering av gener, bland annat gener som gör att hanliga eller honliga könshormoner senare bildas. Hormonerna påverkar könsorganens utveckling och funktion i fostret och senare i livet. De påverkar också de sekundära könskaraktärerna hos vuxna individer. Till de senare hör hos många fåglar bland annat fjäderdräktens färger och kroppens storlek. Könshormonernas roll i fågelfostret är så vitt jag vet oklar.
Hur fungerar miljöbetingad könsbestämning?
Hos de flesta reptiler med temperaturbetingad könsbestämning avgörs könet med en helt annan mekanism. Man vet inte var de receptorer som mäter temperaturen är belägna. Man tror att de finns i genitalkammarna, men man kan inte utesluta möjligheten, att genitalkammarna påverkas av receptorer i andra delar av kroppen via nerver. Hos amerikansk alligator har man dock visat att temperaturreceptorn TRPV4 mäter temperaturen i genitalkammarna. Inom ett visst temperaturintervall tycks denna receptor påverka gener som styr könsbestämningen. Märkligt nog verkar TRPV4 bara styra utvecklingen till hane, inte till hona. Det förefaller således finnas en annan mekanism, som styr den honliga utvecklingen.
Könet betäms hos reptilerna av de honliga hormonerna, östrogenerna. Deras mekanism är således radikalt annorlunda än däggdjurens. Enzymet aromatas tycks spela en central roll, se bilden överst på denna sida. Detta enzym är hastighetsbestämmande i den syntesväg, genom vilken östrogenet estradiol bildas. Om aromataset aktiveras ökar östrogensyntesen och fostret utvecklas till en hona. Om aromatasaktiviteten förblir låg, utvecklas fostret till en hane. Om aromataset hämmas utvecklas hanar även vid temperaturer, som normalt borde leda till honlig utveckling. Mekanismerna är emellertid dåligt utredda och det finns väsentliga skillnader mellan olika arter.
Vad kom först, miljöbetingad eller genetisk könsbestämning?
Det är mycket egendomligt att könet bestäms på så många olika sätt hos ryggradsdjuren genom radikalt olika varianter av genetisk och miljöbetingad könsbestämning. Möjligen kan könet till och med hos vissa arter påverkas av tillväxthastigheten. Hos en nejonögeart uppkommer ett kraftigt överskott av hanar i miljöer med dålig födotlllgång, där larverna växer långsamt. I miljöer med god födotillgång, där larverna växer snabbt, uppkommer även där ett överskott av hanar, men betydligt mindre. Detta kan mycket väl vara funktionellt, eftersom det kräver mer energi att producera ägg än att producera spermier.
Till detta kommer att det finns kräldjursarter vars kön bestäms både av könskromosomer och av temperaturen. Hos en skinkart med könskromosomer har hanarna antingen XX eller XY, honorna alltid XX. XX innebär två X-kromosomer, XY en X-kromosom och en Y-kromosom. Hos denna ödla gör låg temperatur under embryots utveckling att XX-embryon utvecklas till hanar. Hos en annan ödla med könskromosomer har honorna antingen ZZ (två Z-kromosomer) elelr ZW (en Z-kromosom och en W-kromosom), men hanarna har alltid ZZ. Hos denna art gör hög temperatur under embryots utveckling att ZZ-embryon utvecklas till honor.
Vilken var då den ursprungliga formen av könsbestämning hos kräldjur, inklusive fåglar, och däggdjur, två grupper som med mycket stor sannolikhet har en gemensam förfader. Man kan bara spekulera. Det finns argument för båda alternativen.
Referenser
S. M. I. Alam et al.: Did lizards follow unique pathways in sex chromosome evolution? (Genes 9:239, 2018).
J. M. Carré et al.: Exogenous testosterone rapidly increases aggressive behavior in dominant and impulsive men (Biological Psychiatry 82:249-256, 2017).
J. M. Carré and N. A. Olmstead: Neuroscience forefront review. Social neuroendocrinology of human aggression, examining the role of competition-induced testosterone dynamics (Neuroscience 286:171-186, 2015).
P. Celec, D. Ostatníková and J. Hodosy: On the effects of testosterone on brain behavioral functions (Frontiers in Neuroscience 9:12, 2015).
S. M. Gartler et al.: Normal histone modifications on the inactive X chromosome in ICF
and Rett syndrome cells: implications for methyl-CpG binding proteins (BMC Biology 2:21, 2004).
G. C. Hays et al.: Population viability at extreme sex-ratio skews produced by temperaturedependent sex determination (Proceedings of the Royal Society B 284:20162576, 2017).
M. Hines: Sex-related variation in human behavior and the brain (Trends in Cognitive Science 14: 448-456, 2010).
M. Johnson Pokorná and Lukáš Kratochvíl: What was the ancestral sex-determining mechanism in amniote vertebrates? (Biological Reviews 91:1-12, 2016).
N. S. Johnson, W. D. Swink, and T. O. Brenden: Field study suggests that sex determination in sea lamprey is directly influenced by larval growth rate (Proceedings of the Royal Society B
284:20170262, 2017).
H. Merchant-Larios and V. Díaz-Hernández: Environmental sex determination mechanisms in reptiles (Sexual Development 7:95-103, 2013).
N. J. Mitchell and F. J. Janzen: Temperature-dependent sex determination and contemporary climate change (Sexual Development 4:129–140, 2010).
K. T. Moeller: Temperature-dependent sex determination in reptiles (Embryo Project
Encyclopedia, updated 25 February 2015).
R. E. Norman et al.: Trait anxiety moderates the relationship between testosterone responses to competition and aggressive behavior (Adaptive Human Behavior and Physiology 1:312-324, 2015).
D. O’Meally et al.: Are some chromosomes particularly good at sex? Insights
from amniotes (Chromosome Research 20:7-19, 2012).
K. A. Pavlov, D. A. Chistiakov and V. P. Chekhonin: Genetic determinants of aggression and impulsivity in humans (Journal of Applied Genetics 53:61–82,k 2012).
R. Sekido and R. Lovell-Badge: Sex determination involves synergistic action of SRY and SF1 on a specific Sox9 enhancer (Nature 453:930-934, 2008).
E. P. Widmayer, H. Raff, and K. T. Strang: Vander's human physiology (12th ed, McGraw-Hill, 2011).
R. Yatsu et al.: TRPV4 associates environmental temperature and sex determination in the
American alligator (Scientific Reports 5:18581, 2015).
Till början på sidan
Till "Djurfakta"
|


