 |
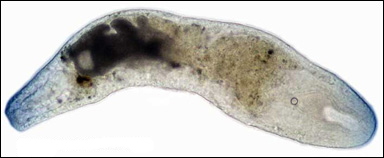
En mycket enkelt uppbyggd mask, ädelstensmasken Proporus bermudensis, som är mindre än 1 millimeter lång. Men den har dock en huvudände (till höger) samt en högersida och en vänstersida. Courtesy of the Turbellarian Taxonomic Database under this CC License. |
|
Kanske såg det första bilateralsymmetriska (tvåsidiga) djuret, en mycket avlägsen förfader till oss, ut ungefär som masken på bilden ovan. Men vi har inga som helst fossil som visar detta enormt viktiga evolutionära framsteg. Kanske hände det så långt tillbaka i tiden som för 800 till 700 miljoner år sedan.
Masken på bilden ovan tillhör stammen Xenacoelomorpha. De flesta av dessa maskar räknades förr till stammen plattmaskar (Platyhelminthes). Numera räknas de alla vanligen som en systergrupp till alla andra bilateralsymmetriska djur (om bilateralsymmetri, se nästa stycke). De placeras därmed vid basen av släktträdet för gruppen Bilateria, de bilateralsymmmetriska djuren. Molekylärbiologiska studier av DNA stödjer starkt denna placering. Det finns dock motsägande studier. Xenacoelomorpha omfattar tre grupper: ädelstensmaskar (Acoela), tvåstensmaskar (Nemertodermatida) och paradoxmaskar (Xenoturbellida).
Fördelar med bilateralsymmetri (tvåsidighet)

Bilateralsymmetriska eller tvåsidiga djur har en huvudände och en bakände, en ryggsida och en buksida samt en högersida och en vänstersida. Genom ett sådant djur kan man bara skära ett enda snitt, som delar djuret i två hälfter, vilka är spegelbilder av varandra. Snittet är lodrät från rygg till buk och från huvud till bakände. Det delar djuret i en högerhalva och en vänsterhalva. Bilateralsymmetri gör det lättare för djur att förflytta sig på ett underlag och att simma. Om de dessutom har mun, hjärna och sinnesorgan organiserade i ett huvud (s.k. cefalisering), blir det ännu lättare för dem att röra sig och också att skaffa sig föda. Texten fortsätter under bilden.
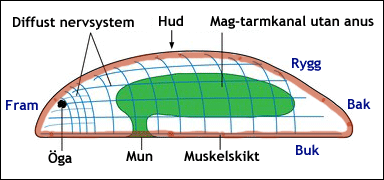 |
Enligt en av flera hypoteser såg det det allra första bilateralsymmetriska djuret ut ungefär så här. Under huden fanns ett eller flera muskelllager. Nervsystemet var bara ett nätverk av nervceller, en hjärna hade ännu inte utvecklats. Kanske fanns det sinnesorgan i framänden, här ett par enkla ögon. Munnen mynnade på buksidan eller i framänden och tarmen slutade blint. Enligt en konkurrerande hypotes var förfadern till Bilateria betydligt mer komplicerat uppbyggd med kroppen uppdelad i segment. I så fall skulle segmenteringen ha bortfallit hos merparten av de nu levande djurstammarna, något som synes mig mindre sannolikt. Courtesy of Nicobola from Wikimedia Commons under this CC Licence. |
|
Bilateralsymmetrin var ett avgörande evolutionärt framsteg. De flesta djur är bilateralsymmetriska. De viktigaste undantagen är svampdjur, plakozoer, nässeldjur (bl.a. maneter och koralldjur) samt kammaneter. Läs mer om bilateralsymmetri på en annan sida.
De enklaste bilateralsymmetriska djurens byggnad
Xenacoelomorpha är de enklast uppbyggda bilateralsymmetriska djuren. De har de tre cellskikt, groddblad, som bygger upp kroppen hos Bilateria: ektoderm, mesoderm och endoderm. Ektodermet bildar hos bilaterier bland annat hud och nervsystem, endodermet mag-tarmkanalen och mesodermet flera vävnader och organ mellan de två andra skikten. Mesodermet saknas hos de ursprungligare djurstammar som står närmare basen för alla djurs släktträd: svampdjur, plakozoer, nässeldjur och kammaneter. Många menar dock att kammaneterna har mesoderm. Alla bilateralsymmetriska djur har mesoderm. Texten fortsätter under bilderna.
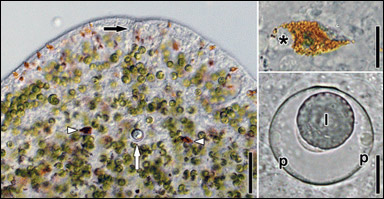 |
Till vänster ses huvudet hos ädelstensmasken Symsagittifera roscoffensis. Det första huvudet, ett enormt evolutionärt framsteg, såg kanske ut ungefär så här. Djuret har i huvudet en ansamling av nervceller, en enkel hjärna, som dock inte syns på bilderna. På den vänstra bilden ser man vad som sannolikt är ett par mycket enkla ögon (vita pilspetsar) och ett balansorgan, en så kallad statocyst (vit pil). Mynningen till en slemkörtel skymtar längst fram (svart pil). Överst till höger ses ett öga i närbild. Det består av en enda cell med bruna ljusbrytande korn. Nederst till höger ses statocysten i närbild med ett omgivande hölje (p) och inuti det en statolit (l). Statoliten påverkas av tyngdkraften och hamnar därför alltid i botten av statocysten. Om masken vänder på kroppen kommer statoliten att flytta på sig och pressas mot en annan del av statocystens hölje. På något sätt, man vet inte hur, kan masken då avläsa kroppens läge i förhållande till tyngdkraften. Courtesy and copyright of J. G. Achatz et al. |
|
Xenacoelomorpha har förvisso de tre groddbladen, men organen är ofta inte välutvecklade och ofta ganska få. De har en framände, men den kan knappast alltid kallas huvud, se dock Symsagittifera i bilden ovan. I Proporus framände, till höger på bilden överst på sidan, ses svalget vars mynning fungerar som både mun och anus, men denna mask har inget välutvecklat huvud. I övrigt saknar djuret hålrum i kroppen. I stället för mag-tarmkanal har Proporus en central massa av sammansmälta endodermala celler, som tar hand om födan. Vissa andra maskar inom gruppen Xenacoelomorpha har en enkelt uppbyggd, blint slutande tarm.
Utsöndringsorgan, motsvarande våra njurar, saknas hos Xenacoelomorpha, men finns hos alla andra Bilateria, i sin enklaste form som så kallade protonefridier. Xenacoelomorpha har en ektodermal hud. Deras nervsystem består av nätverk av nervceller i kroppen särskilt i anslutning till huden. En del av dem har också framtill en klunga av nervceller, som utgör en enkel hjärna. Gruppen har även mesodermala muskelceller bland annat under huden och runt svalget. Musklerna kan ändra kroppens form. De rör sig dock huvudsakligen med hjälp av flimmerhår (cilier) på bukens hudceller. Könsorganens har en mycket varierande uppbyggnad.
Gener som styr embryots utveckling
I en studie från 2018 visade man att genomet hos Xenacoelomorpha innehåller alla de elva genklasser av så kallade homeotiska gener, som styr embryonalutvecklingen och därmed kroppens byggnadsplan hos bilaterier. Man antog att dessa genklasser fanns redan hos bilateriernas förfader. I så fall skulle den verktygslåda, som möjliggör utveckling av nya djurgrupper med radikalt annorlunda byggnadsplaner funnits långt innan den så kallade kambriska revolutionen för cirka 500 miljoner år sedan. Då uppträdde i ett geologiskt perspektiv mycket plötsligt de flesta nutida djurstammar på den fossila arenan. Detta stöder tanken att dessa djurstammar hade en lång evolutionär historia innan den kambriska revolutionen. Men man har hittat mycket få eller inga fossil från prekambrisk tid, delvis kanske beroende på att djuren ännu inte utvecklat skal och skelett, hårda strukturer som lätt bevaras som fossil.
Avvikande tolkningar
Enligt en avvikande åsikt är Xenacoelomorpha inte ursprunglig. I så fall har den förlorat en rad anatomiska strukturer och härstammar från djur med mera komplicerad kroppsbyggnad. Släktskapen mellan de tre undergrupperna inom Xenacoelomorpha har också ifrågasatts.
Referenser
J. G. Achatz et al.: The Acoela: on their kind and kinships, especially with nemertodermatids and xenoturbellids (Bilateria incertae sedis) (Organisms, Diversity & Evolution 3:267-286, 2013).
Jaume Baguñá et al.: Back in time: a new systematic proposal for the Bilateria (Philosophical Transactions of the Royal Society B 363:1481-1491, 2008).
M. Brauchle et al.: Xenacoelomorpha survey reveals that all 11 animal
homeobox gene classes were present in the first bilaterians (Genome Biology and Evolution 10:2205-2217, 2018).
J. T. Cannon et al.: Xenacoelomorpha is the sister group to Nephrozoa (Nature 530:89-93, 2016).
Brenda Gavilán, E. Perea-Atienza, and P. Marínez: Xenacoelomorpha: a case of independent nervous system centralization? (Philosophical Transactions of the Royal Society B 371:20150039, 2016).
U. Jondelius et al.: How the worm got its pharynx: phylogeny, classification and Bayesian assessment of character evolution in Acoela (Systematic Biology 60:845-871, 2011).
U. Jondelius et al.: The Nemertodermatida are basal bilaterians and not members of the Platyhelminthes (Zoologica Scripta 31:201-215, 2002).
H. Nakano et al.: A new species of Xenoturbella from the western Pacific Ocean and the evolution of Xenoturbella (BMC Evolutionary Biology 17:245, 2017).
C. Nielsen: Animal evolution, interrelationships of the living phyla (3rd ed, Oxford University Press, 2012).
H. Philippe et al.: Acoelomorph flatworms are deuterostomes related to Xenoturbella (Nature 470:255-258, 2011).
Till början på sidan
Till "Djurfakta"
|


