 |
Embryots utveckling speglar evolutionen, men bara delvis

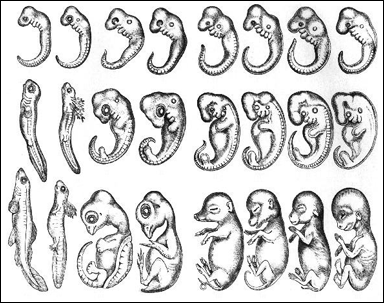
Denna bild från 1800-talet visar embryots utveckling hos ett antal ryggradsdjur, från fisk (till vänster) till människa (till höger). De tidigaste stadierna är överst. Det i texten nedan diskuterade fylotypiska stadiet motsvaras av den översta raden. Bilden illustrerar Ernst Haeckels så kallade biogenetiska regel. Denna regel innebär att embryots utveckling alltid helt och hållet återspeglar ett djurs evolution. Biologer har sedan mer än hundra år vetat att bilden är i flera avseenden felaktig, eventuellt förfalskad. Haeckel erkände detta redan 1891. Biologer kom också för många år sedan fram till att den biogenetiska regeln i sin strikta form är felaktig. Trots detta används fortfarande den felaktiga bilden och den icke korrekta biogenetiska regeln av företrädare för kreationism och "intelligent design" som ett argument mot evolutionen. Den biogenetiska regeln är förvisso fel, så som den formulerades av Haeckel. Numera vet man dock att vissa embryonala strukturer återspeglar ett djurs evolution. Men man vet också att andra embryonala strukturer, under inflytande av evolutionens naturliga urval, undergått så stora förändringar att de inte längre återspeglar djurets evolution. För en utförlig diskussion, se texten nedan. From G. J. Romanes after E. Haeckel, in the public domain. |
|
Vissa strukturer som funnits hos våra evolutionära förfäder kan anläggas i embryot under fostertiden, för att sedan försvinna. Valfoster anlägger bakbensknoppar som sedan försvinner. Fungerande gälspringor och gälbågar fanns hos våra förfäder bland fiskarna. Men gälfickor och gälbågar bildas även hos foster av människor och andra fyrfotadjur. Gälfickorna förbinds dock inte med svalget. Gälspringor bildas således inte. Gälfickorna och gälbågarna finns kvar under en period, men försvinner sedan.
Flera strukturer i huvudregionen härstammar dock från gälbågarna, hos däggdjuren bland annat mellanörat med dess ben (hammaren, städet och stigbygeln), bisköldkörtlarna, brässen (thymus), tungbenet (hyoidbenet), delar av struphuvudet samt delar av ansiktsmuskulaturen.
Det är mycket viktigt att framhålla att embryots utveckling inte strikt återspeglar ett djurs evolution. Alla organ och strukturer hos de evolutionära förfäderna återfinns således inte i ett embryo. Men man kan ändå belysa vissa strukturers evolution genom att studera deras utveckling i embryot. Detta forskningsområde kallas numera "evo-devo".
Placentadäggdjur, pungdjur och kloakdjur
Placentadäggdjurens, inklusive människans, tidiga embryonalutveckling är ett intressant exempel på att vissa stadier återspeglar evolutionen, medan andra inte gör det. Placentadäggdjur (moderkaksdäggdjur, äkta däggdjur) och pungdjur föder levande ungar, som får näring av modern via en moderkaka (placenta). Men båda grupperna härstammar från äggläggande djur. Kloakdjuren (näbbdjuret och myrpiggsvinen) är däggdjur som fortfarande lägger ägg. Låt oss jämföra de tidiga embryonas utveckling hos placentadäggdjuren med utvecklingen hos de äggläggande släktingarna kräldjur, inklusive fåglar, och kloakdjur.
Embryots tidiga utveckling hos djur
Hos alla djur börjar den befruktade äggcellens utveckling med celldelningar som kallas klyvningsdelningar. Hos de flesta djurgrupper delas då hela äggcellen upp i flera celler, så att en klump, morula, bestående av flera celler bildas. Morulan ger upphov till en vanligen ihålig blastula. Blastulan utvecklas till en tvåskiktad gastrula med en öppning, urmunnen. Läs om djurs tidiga embryonalutveckling på en annan sida.
Reptilerna, inklusive fåglarna, och kloakdjuren är några av flera undantag bland djuren. Hos dem klyvs inte ägget fullständigt. Endast ett litet område på äggets yta delas upp i celler och bildar en så kallad groddskiva. Detta beror på att dessa ägg har ett extremt stort förråd av upplagsnäring, gula, i form av så kallade gulekorn. Det är endast groddskivan som ger upphov till embryot. Det är bara embryot som utvecklas till den unge som kläcks ur ägget. Texten fortsätter under faktarutan.
|
|
|
Människans tidiga embryonala utveckling
Den tidiga utvecklingen hos ett människoembryo fram till dess att groddskivan bildas. Du kan lyssna till videons engelska speakertext eller läsa följande utförliga beskrivning på svenska.
Efter ett dygn (Day +1) har befruktningen skett och man ser två cellkärnor (pronuclei) inuti äggcellen, en hanlig (från en spermie) och en honlig (tillhörande äggcellen). Båda har ett halverat kromomantal. Äggcellen kallas nu zygot. Nedtill utanför zygoten syns en polkropp, som bildats vid den blivande äggcellens meios (reduktionsdelning). Meiosen omfattar två celldelningar. Kromosomtalet halveras under den första av dessa. Äggcellen behåller emellertid nästan all sin cytoplasma. De andra dottercellerna kallas polkroppar. De är mycket små och belägna utanför äggcellens membran.
Under det andra dygnet (Day +2) smälter de båda pronuclei ihop samtidigt som zygoten delar sig och ger upphov till ett embryo med två celler. Dessa celler har det normala kromosomantalet i sina kärnor, med hälften av kromosomerna från fadern och hälften från modern. Embryot genomgår sedan två celldelningar till (Day +2, Day +3) och består nu av åtta celler. Efter ytterligare celldelningar (Day +4) bildar det en cellklump som kallas morula.
Morulan omvandlas efter ännu fler celldelningar till en klotformad, ihålig blastocyst (Day +5). Det cellskikt som omger den kallas trofoblast och ger så småningom upphov till den yttersta fosterhinnan, korion. Innanför trofoblasten (upptill till vänster på videon) finns en cellklump, den inre cellmassan, som senare bildar en groddskiva. Det egentliga embryot bildas i groddskivan och utvecklas så småningom till ett foster. Groddskivan utvecklas i fortsättningen på i princip samma sätt hos däggdjur som hos reptiler, inklusive fåglar. Läs om fosterhinnorna och om ryggsträngen på andra sidor. Förloppen visas betydligt snabbare än de verkligen sker. From YouTube, courtesy of Atlas de Reproducción Asistida. |
|
Placentadäggdjurens embryonala utveckling
Hos placentadäggdjuren försörjs ju embryot med näring från modern via moderkakan (placentan). Då behövs inte så mycket gula. Äggen har därför förlorat större delen av gulan och är gulefattiga. Märkligt nog har de i samband med detta återgått till den ursprungliga typen av klyvningsdelningar. Hela ägget delas upp i celler så att en morula uppstår. Men om man studerar den fortsatta utvecklingen, framstår det ändå som fullständigt klart, att vi placentadäggdjur härstammar från äggläggande förfäder med gulerika ägg. Den så kallade inre cellmassan i embryot delas hos oss upp i två skikt, groddblad. Det övre ektodermet ger bland annat upphov till överhud och nervsystem, det undre endodermet ger bland annat upphov till magtarmkanalens innersta cellskikt och dess körtlar. Här finns stora likheter med utvecklingen hos de äggläggande släktingarna. Ännu större likheter finner man i nästa steg, gastrulationen. Precis som hos de äggläggande släktingarna bildas en så kallad primitivstrimma genom vilken celler vandrar in mellan ektoderm och endoderm och bildar det tredje groddbladet, mesodermet.
De följande stegen i utvecklingen sker på likartat sätt hos alla ryggradsdjur. Mesodermet bildar i embryots mittlinje den så kallade ryggsträngen (kordan). På båda sidor om ryggsträngen bildas rader av somiter. Somiterna är klossar av mesodermceller. De ger bland annat upphov till läderhud och skelettmuskulator. Senare snörps ett nervrör av från ektodermet på ryggsidan ovanför ryggsträngen. De inre delarna av somiterna utvecklas till ryggkotor, som omsluter kordan och nervröret. Läs om nervröret på en annan sida. Texten fortsätter under videon.
|
Videon visar primitivstrimman på ryggsidan i ett tidigt kycklingembryo, se texten ovan. Embryots framände ligger överst. Man ser ytan på ektodermet, det yttersta cellskiktet. Ektodermcellerna (vita fläckar) vandrar mot strimman. Där rör de sig neråt och omvandlas till mesoderm. Mesodermcellerna sprider sig utåt från primitivstrimman under ektodermet, det vill säga i utrymmet mellan ektodermet och det under detta belägna endodermet. De finns då i det vita område som blir synligt runt strimman i slutet av videon. Mesodermet bildas på samma sätt i ett människoembryo. Förloppet visas betydligt snabbare än det verkligen sker. From YouTube, courtesy of Vincent Fleury, Directeur de recherches au CNRS, HDR. |
|
Placentadäggdjurens förfäder hade gulerika ägg
Primitivstrimmorna i ett människoembryo och ett hönsembryo ser nästan precis likadana ut och fungerar på samma sätt. Detta är ett starkt argument för att placentadäggdjuren och pungdjuren härstammar från djur med gulerika ägg och groddskiva. Ett andra starkt argument är att placentadäggdjuren och pungdjuren utvecklar precis samma fosterhinnor som kräldjur, inklusive fåglar, och kloakdjur. Läs om däggdjurens fosterhinnor på en annan sida. Ett tredje starkt argument är naturligtvis att kloakdjuren lägger gulerika ägg med groddskiva, men ändå har en rad egenskaper gemensamma med övriga däggdjur, till exempel hår och mjölkkörtlar. Och ett fjärde starkt argument är att icke fungerande rester av de gener som kodar för vitellogeninerna finns hos både pungdjur och placentadäggdjur. Vitellogeninerna är lipoproteiner och utgör den viktigaste komponenten i gulans näringsförråd hos de äggläggande reptilerna, inklusive fåglarna. Till yttermera visso har näbbdjuret, ett äggläggande kloakdjur, behållit en fungerande vitellogeningen. Texten fortsätter under videon.
|
Videon visar hur primitivstrimman i ett tidigt kycklingembryo slutes och försvinner med början i dess framände, den så kallade Hensens nod (till vänster). Detta sker efter det att mesodermet bildats. Förloppet visas betydligt snabbare än det verkligen sker. From YouTube, courtesy of Vincent Fleury, Directeur de recherches au CNRS, HDR. |
|
Det fylotypiska stadiet och timglasmodellen
Det så kallade fylotypiska stadiet är en period under ryggradsdjurens embryonalutveckling, då de uppvisar stora likheter. Detta stadium infaller efter nervrörsbildningen. Embryot är då uppdelat i segment från framänden till bakänden och flera olika organ har bildats. Stadiet kallas också faryngula, eftersom gälfickor och gälbågar då har utvecklats i svalget (pharynx).
Det är mycket viktigt att notera att faryngulorna hos olika ryggradsdjur inte ser exakt likadana ut. En berömd bild publicerad av Ernst Haeckel och andra på 1800-talet är i flera avseenden felaktig och visar bland annat alltför stora likheter mellan olika djurs embryon. Biologer är sedan länge medvetna om detta, se bildtexten överst på sidan. Icke desto mindre gör kreationister fortfarande ett stort nummer av denna felaktiga bild som publicerades för mer än hundra år sedan. Texten fortsätter under videon.
|
Video visar hu nervröret slutes i ett kycklingembryo, med början i framänden (till höger). Det ser ut som när ett blixtlås låses. Nervröret ger upphov till hjärna och ryggmärg. Samtidigt bildas allt fler somiter (vita fyrkanter) på båda sidor om nervröret. Förloppet visas betydligt snabbare än det verkligen sker. From YouTube, courtesy of Boryskina-Al-Kilani-Fleury/CNRS/MSC. |
|
Enligt den så kallade timglasmodellen finns det restriktioner under det fylotypiska stadiet, alltså hos faryngulan, som gör att alla ryggradsdjursembryon då måste vara uppbyggda efter en liknande grundplan. Utvecklingen före och efter det fylotypiska stadiet är mer flexibel. Därför finns det stora skillnader mellan olika grupper av ryggradsdjur under deras tidiga och deras sena utvecklingsstadier. Det fylotypiska stadiet motsvarar timglasets insnörning. Ett exempel är de stora skillnaderna mellan den tidiga utvecklingen hos däggdjuren och hos andra fyrfotadjur som beskrivits ovan. Den senare utvecklingen, efter faryngulastadiet, skiljer sig ju också åt hos olika ryggradsdjur. Slutresultaten blir, som vi alla vet, mycket olika varandra: en fisk, en fågel eller ett däggdjur.
Timglasmodellen stöds således av de iakttagelser som man kunnat göra av embryonas utveckling hos olika ryggradsdjur. Intressant nog stöds modellen också av molekylärbiologiska studier av genaktiviteterna under embryonas utveckling. Genuttrycksmönstret hos olika djur är mycket likartat under det fylotypiska stadiet. Detta innebär att det är i stort sett samma gener som är aktiva hos de olika djuren. Skillnaderna i genaktiviteter mellan olika djurgrupper är mycket större före och efter det fylotypiska stadiet.
Vad är det då som begränsar utrymmet för förändringar under det fylotypiska stadiet? Detta är fortfarande oklart, men det finns flera hypoteser. Enligt en hypotes är det nätverk av signaler, som styr utvecklingen under det fylotypiska stadiet, så komplicerat, att även små förändringar leder till kraftiga störningar. En förändrad signal kan till exempel leda till effekter i flera olika delar av embryot. Signalerna förmedlas bland annat av proteinfaktorer. Enligt en annan hypotes måste de så kallade Hox-generna uttryckas efter ett koordinerat mönster under det fylotypiska stadiet, annars utvecklas kroppens grundplan inte på ett normalt sätt. Läs om hox-generna på en annan sida.
Referenser
D. Brawand, W. Wahli and H. Kaessmann: Loss of egg yolk genes in mammals and the origin of lactation and placentation (PLoS Biology 6: e63, 2008).
A. Collazo: Developmental variation, homology, and the pharyngula stage (Systematic Biology 49:3–18, 2000).
N. Irie and S. Kuratani: Comparative transcriptome analysis reveals vertebrate phylotypic period during organogenesis (Nature Communications 2, Article number: 248, doi:10.1038/ncomms1248, 2011).
A. T. Kalinka et al.: Gene expression divergence recapitulates the developmental hourglass model (Nature 468:811-814, 2010).
K. V. Kardong: The vertebrates (5th ed, McGraw-Hill, 2009).
B. Piaseckaet al.: The hourglass and the early conservation models - co-existing patterns of developmental constraints in vertebrates (PLoS Genetics 9: e1003476, 2013).
M. K. Richardson and G. Keuck: Haeckel's ABC of evolution and development (Biological Reviews of the Cambridge Philosophical Society 77:495-528, 2002).
W. Shumway and F. B. Adamstone: Vertebrate embryology (5th ed, Wiley, 1962).
F. Wilt and S. Hake: Principles of developmental biology (W.W. Norton & Company, 2004).
Till början på sidan
Till "Djurfakta"
|


