 |
 |
Överst syns ett försök till rekonstruktion av Pikaia gracilens, ett djur som troligen stod nära den gemensamma förfadern till alla djur inom djurstammen ryggsträngsdjur (Chordata). Ryggradsdjuren, inklusive människan, är en understam bland ryggsträngsdjuren. Vi härstammar sannolikt inte från Pikaia, men Pikaia är det äldsta kända fossil som kan anknytas ryggsträngsdjurens urfader och därmed till människans stamträd. Pikaia levde under kambriumperioden för cirka 505 miljoner år sedan. Notera att muskulaturen utgörs av en rad v-formade skikt, från framänden med sina tentakler (till vänster) till bakänden med sin stjärtfena (till höger). Likheten med de v- eller w-formade muskelskikten hos de flesta av dagens fiskar är slående.
Underst ses en lansettfisk (understammen Cephalochordata) med huvudet till vänster. Molekylärbiologiska data tyder på att lansettfiskarna är den nu levande djurgrupp som står närmast ryggsträngsdjurens urfader. Lansettfiskarna uppvisar dessutom stora anatomiska likheter med Pikaia, läs mer om detta i texten nedan. From Encyclopedia of Life, courtesy of Muséum national d'histoire naturelle and Citron under this CC License (above) and Gustav Paulay under this CC License (below). |
|
Vilken är människans äldsta kända förfader? Vilket nu levande djur är mest likt denna förfader? De äldsta fossil som vi känner till i vår utvecklingshistoria är cirka 500 miljoner år gamla. Fantastiskt nog visar det sig att dessa fossil är mycket lika en nu levande djurgrupp, lansettfiskarna, så till den grad att lansettfiskarna kan betraktas som levande fossil. De är, trots sitt namn, inte fiskar.
Människan är ett däggdjur. Däggdjuren tillhör understammen ryggradsdjur (Vertebrata) inom stammen ryggsträngsdjur (Chordata) bland djuren. Till ryggradsdjuren hör alla fiskar och alla fyrfotadjur (d.v.s. groddjur, kräldjur inklusive fåglar, och däggdjur). Ryggsträngsdjuren inkluderar ytterligare två understammar: manteldjur (Urochordata; se bilden nedan) och lansettfiskar (Cephalochordata, se bilden ovan). Manteldjuren och lansettfiskarna har en betydligt enklare kroppsbyggnad än ryggradsdjuren. De har därför sedan gammalt ansetts stå närmare ryggsträngsdjurens urfader än ryggradsdjuren. Detta har nu bekräftats av molekylärbiologiska studier av arvsmassans DNA.
Ryggsträngsdjurens urfader levde förmodligen långt innan den så kallade "kambriska explosionen", som inleddes för cirka 540 miljoner år sedan. Notera att Pikaia, det ursprungligaste kända fossila ryggsträngsdjuret, levde betydligt senare, för cirka 505 miljoner år sedan. Se bilden överst på sidan. Under den kambriska explosionen uppträdde plötsligt de flesta av de artrikaste nu levande djurstammarna på den fossila arenan. Orsakerna till detta är oklara. Det finns emellertid ett flertal hypoteser. Molekylärbiologiska data tyder på att det kanske inte var någon explosion. I stället uppkom kanske flera av djurgrupperna långt innan kambriumperioden började. Orsaken till att man har hittat så få fossil från tiden före kambrium skulle kunna vara djuren då saknade mineraliserade skelett och skal. Mjukdelar bevaras mycket sällan som fossil.
Lansettfiskarna är de nutida djur som står närmast ryggsträngsdjurens urfader, men de är inte ryggradsdjurens närmaste nu levande släktingar. Förvånande nog tyder DNA-studier i stället på att det är manteldjuren som står oss ryggradsdjur närmast. Man kan ju knappast tänka sig ett djur som är mer olikt ett ryggradsdjur än manteldjuret på den översta bilden nedan, en vuxen sjöpung. Förklaringen är att manteldjurens larver ser helt annorlunda ut än de vuxna djuren och har en rad anatomiska egenskaper som är gemensamma för alla ryggsträngsdjur. Mer om dessa egenskaper senare. Som synes på den nedersta bilden nedan, påminner manteldjurens larver till det yttre om grodlarver. De genomgår en drastisk omvandling, en metamorfos, som resulterar i ett vuxet djur med en radikalt annorlunda kroppsbyggnad. En grupp bland manteldjuren behåller dock flera larvala egenskaper som vuxna. Texten fortsätter under bilderna.
 |
 |
Överst syns ett exemplar av ett vuxet manteldjur, sjöpungen Ciona intestinalis. De intar sin föda genom att filtrera havsvattnet och tillvarata plankton och organiska partiklar. Vattnet strömmar in genom den ena öppningen och ut genom den andra. Födan fastnar i gältarmen. Mer om den längre ner på denna sida.
Nederst ses en larv av ett manteldjur. Som synes är den mycket olik de vuxna djuren. Läs mer om detta i texten ovan. Courtesy of Bernard Picton (above) and Arjan Gittenberger (below) from Encyclopedia of Life under this CC License. |
|

Genomfördubblingar och Hox-gener
Lansettfiskarna och manteldjuren har en mycket enkelt uppbyggd kropp. Det samma gällde Pikaia, det ursprungligaste kända fossila ryggsträngsdjuret, som levde för cirka 505 miljoner år sedan. Se bilden överst på sidan. Ryggradsdjurens förfader påminde med största sannolikhet om Pikaia och lansettfiskarna. Hur kunde ett så enkelt uppbyggt djur, utan hjärna och hjärta, ha gett upphov till den utvecklingslinje, som ledde fram till dagens komplicerat uppbyggda ryggradsdjur, bland dem människan? Förklaringen finner man i en genomgripande händelse som ägde rum för mer än 500 miljoner år sedan. Vid två tillfällen fördubblades hela genomet (arvsmassan) hos den utvecklingslinje bland ryggsträngsdjuren, som senare gav upphov till ryggradsdjuren. Man tror numera att detta skedde hos en urfader till alla ryggradsdjur. Men den andra genomfördubblingen kan också ha skett efter det att de käklösa ryggradsdjuren, rundmunnarna (Agnatha), grenades av från släktträdet, men innan de käkförsedda ryggradsdjuren (Gnathostomata) avgrenades. Till rundmunnarna hör i dag bara pirålar och nejonögon. Alla övriga ryggradsdjur är käkförsedda. De inkluderar alla fiskar, utom pirålar och nejonögon, samt alla fyrfotadjur, det vill säga groddjur, kräldjur inklusive fåglar och däggdjur. Om två genomfördubblingar inte hade skett, så hade det inte funnits några ryggradsdjur på jorden, utom möjligen pirålar och nejonögon med en genomfördubbling. Texten fortsätter under faktarutan.
|
|
|
Världens slemmigaste slem
Den översta videon visar en pirål som producerar slem. Pirålarna (Myxini) anses stå närmast ryggradsdjurens ursprung bland de nu levande ryggradsdjuren. Till deras primitiva drag hör avsaknaden av såväl ryggrad (!) som käkar och pariga fenor. Men de har också flera intressanta specialiseringar. De utsöndrar ett unikt slem från körtlar som ligger i rad längs med kroppssidorna. Detta tvåkomponentslem är djurvärldens slemmigaste slem. Det består dels av så kallade muciner som kan absorbera enorma mängder vatten och som gör slemmet mycket trögflytande, dels av trådformade proteiner som bildar ett nätverk och därmed gör slemmet utomordentligt segt. Slemmet aktiveras först när det kommer ut i vattnet. Inuti körtlarna är mucinerna förpackade i små blåsor och proteinerna hopslingrade. Slemmet fungerar med största sannolikhet som ett försvar mot rovfiskar och andra predatorer, som kan kvävas när slemmet fastnar i deras gälar. Den nedre videon visar hur flera rovfiskar hugger tag i pirålar, men genast släpper bytet, avskräckta av slemmet. Det är en fascinerande spekulation att dessa ursprungliga djur överlevt i hundratals miljoner år tack vare sitt slem. Läs om hur pirålarna kan slå knut på sig själva på en annan sida. From YouTube, courtesy of the Vancouver Aquarium (above) and Museum of New Zealand Te Papa Tongarewa (below). |
|
Två genomfördubblingar ledde till att det i princip fanns fyra gener i stället för en av alla genomets gener, egentligen fyra par av varje gen fördelade på fyra kromosompar. Nästan alla kroppscellernas gener hos djur finns nämligen normalt i två exemplar, ett på vardera kromosomen i ett kromosompar. Det fanns nu tre nya genkopior, som inte var nödvändiga. De flesta av de nya genkopiorna hos urryggradsdjuren försvann emellertid så småningom från genomet. Andra drabbades av skadliga mutationer, som ledde till att de inte fungerade längre. De blev då genetisk barlast, så kallade pseudogener. Men många nya genkopior fortsatte att fungera. Man kan fråga sig hur avkomlingar, försedda med ett ökat antal identiska genkopior, kunde utveckla en kropp med ett mera komplicerat byggnadsplan. Förklaringen är att många av de nya genkopiorna under inflytande av naturligt urval genomgick en evolution så att de fick förändrade funktioner jämfört med den ursprungliga genen. Särskilt betydelse hade detta för de så kallade homeotiska generna, som styr uppkomsten av kroppens byggnadsplan under embryots utveckling. De mest kända bland de homeotiska generna är Hox-generna, som hos djur styr utvecklingen av kroppens delar i dess längsriktning, hos däggdjur från huvudet till svansen. Hox-generna ligger vanligen intill varandra längs med en kromosom i samma ordningsföljd som de kroppsdelar de påverkar. Se bilden nedan. Texten fortsätter under bilden.
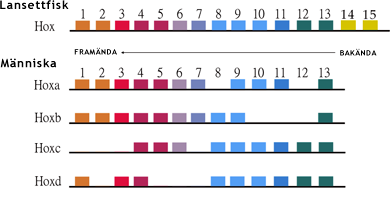 |
Lansettfiskarna har 15 Hox-gener (överst). Efter två genomfördubblingar erhöll ryggradsdjuren fyra uppsättningar Hox-gener. Förlust av flera Hox-gener har emellertid lett till att däggdjuren och de flesta andra ryggradsdjursgrupper har 39 Hox-gener, inte sextio (nederst). Detta har bidragit till att ryggradsdjuren kunnat utvecklat en betydligt mer komplicerad byggnadsplan för kroppen än lansettfiskarna. Hox-generna kodar för så kallade transkriptionsfaktorer, proteiner som aktiverar eller inaktiverar andra gener. Dessa geners proteinprodukter styr i sin tur utvecklingen av kroppens olika delar. Hox-gener förekommer hos alla djur, utom ett fåtal enkelt byggda djurstammar, till exempel svampdjuren. Modified image. Courtesy of Bstlee, in the public domain. |
|
Lansettfiskarna har en uppsättning Hox-gener. Ryggradsdjuren har fyra uppsättningar Hox-gener. De har emellertid förlorat några Hox-gener. Därför har de inte fyra gånger fler Hox-gener än lansettfiskarna. Men de har avsevärt fler Hox-gener med olika funktioner än lansettfiskarna. De har också fler homeotiska gener med andra funktioner än Hox-generna. Ett ökat antal homeotiska gener med nya funktioner är den sannolikt viktigaste orsaken till att ryggradsdjuren har kunnat utveckla en avsevärt mera komplicerad kroppsbyggnad än lansettfiskarna och även flera nya organ.
Ryggsträngsdjurens utveckling
Ryggsträngsdjuren förenas av ett antal gemensamma anatomiska strukturer. Hos många av dem uppträder dessa strukturer bara under embryots utveckling i ägget, hos en del också under larvstadiet, men de finns där.
Ryggsträngen (kordan) och ryggraden
Ryggsträngen är den struktur som gett namn åt djurgruppen. Den kallas även korda. Kordan är en elastisk stav som löper längs med ryggen och fungerar som skelett. Den innehåller en kärna av celler och utanför dem vätska, ibland finns vätskan inuti cellerna. Utanför kärnan finns ett segt hölje av bindvävnad. Eftersom vätskan inte kan pressas ihop, kan kordan inte förkortas. När muskulaturen på ena sidan av kroppen dras ihop, böjs I stället kordan, och därmed hela kroppen, i en båge. När muskulaturen slappnar av fjädrar den elastiska kordan tillbaka, så att kroppen åter blir rak. Detta gör att djuret kan simma på ett effektivt sätt. Kordan fungerar som ett så kallat hydrostatiskt skelett. Se bilden nedan. Texten fortsätter under bilderna.
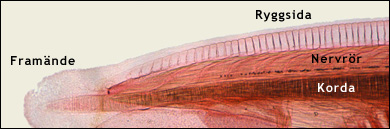 |
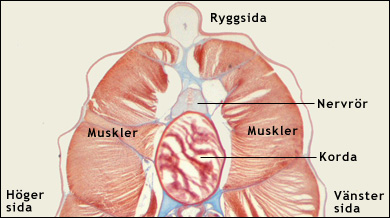 |
Överst ses övre delen av framänden av en lansettfisk. På ryggsidan ser man kordan och ovanför den nervröret.
Nederst ses överdelen av ett tvärsnitt genom en lansettfisk, bakom framänden. Man ser kordan med dess vätskerika kärna och dess bindvävshölje, nervröret med ett svagt urskiljbart hålrum samt muskulaturen på båda sidorna av kroppen. Läs mer i texten ovan och nedan. Färgade ljusmikroskopiska snitt. Modified images. Courtesy of John Houseman, from Wikimedia Commons under this GNU License. |
|
En välutvecklad korda finns hos de vuxna lansettfiskarna och hos manteldjurens larver. De flesta manteldjur saknar korda som vuxna. Ryggradsdjuren kan, som ovan nämnts, indelas i två grupper. Till rundmunnarna (Agnatha) hör pirålar och nejonögon. De har inte utvecklat käkar. De övriga ryggradsdjuren (Gnathostomata) har käkar. Rundmunnarna har en välutvecklad korda även som vuxna. Hos pirålarna utgörs ryggskelettet enbart av en korda. Nejonögonen har dessutom ryggkotor. Hos de käkförsedda ryggradsdjuren (Gnathostomata) har ryggraden med sina kotor av brosk eller benvävnad i regel tagit över kordans funktioner. Kordan finns dock kvar hos en del vuxna fiskar, bland annat hos lungfiskar och störfiskar. Hos människan utgör den geléartade kärnan i de mellan ryggkotorna belägna diskarna en rest av kordan. Flera anatomiska egenheter hos pirålarna, bland annat frånvaron av ryggrad, gör att man ibland räknar dem som en systergrupp till ryggradsdjuren inom den överordnade gruppen Craniata ("djur med kranier"). Denna indelning stöds emellertid inte av molekylärbiologiska data.
Nervröret och nervsystemet
Ett andra kännetecken för ryggsträngsdjuren är nervsystemet anläggs längs med embryots hela ryggsida som en långsträckt inbuktning av ektodermet, embryots yttersta cellskikt, som också ger upphov till överhuden. Inbuktningen fördjupas och snörps av från huden som ett nervrör. Hos vuxna lansettfiskar finns nervröret kvar med en uppsvällning i framändan som tros vara ett förstadium till en hjärna. Hos ryggradsdjuren omvandlas det enkla nervröret under embryots utveckling till deras komplicerade nervsystem med hjärna och ryggmärg. Se bilden ovan. På en annan sida finns en bild som visar hur nervröret anläggs i embryot. Texten fortsätter under bilderna.
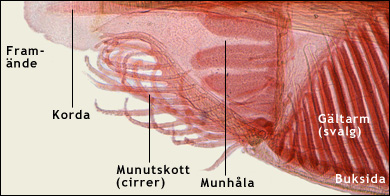 |
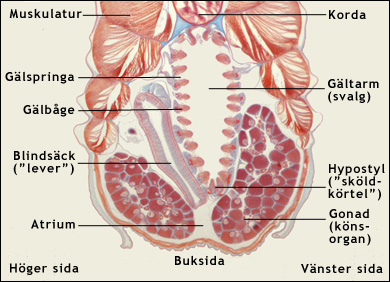 |
Överst ses underdelen av framänden av en lansettfisk. Man ser munhålan och den komplicerade munapparaten, som bland är försedd med munutskott (cirrer). Bakom munhålan finns svalget som är utformat som en gältarm med gälbågar och mellan dem gälspringor.
Nederst ses undre delen av ett tvärsnitt genom en lansettfisk, bakom framänden. Man ser gältarmen med en mängd gälbågar och mellan dem öppningar, gälspringor. Precis som manteldjuren, intar lansettfiskarna sin föda genom att filtrera bort plankton och organiska partiklar från havsvattnet. Vattnet tas in genom munnen, varvid grövre partiklar filtreras bort av munapparaten. Det passerar sedan ut genom gälspringorna. Då fastnar ätbara partiklar på de slemklädda gälbågarna. Slemmet med de matnyttiga partiklarna transporteras sedan av cellernas flimmerhår (cilier) uppåt längs med gälbågarna och sedan bakåt längs med en fåra i gältarmens tak. Till slut når slemmet den egentliga tarmen, bakom gältarmen. Där bryts födan ner och nedbrytningsprodukterna absorberas till blodet. Vattnet som lämnar gältarmen hamnar i atrium, ett särskilt hålrum som släpper ut vattnet genom en atriopor på djurets undersida.
I gältarmens botten finns hypostylen, som är en föregångare till ryggradsdjurens sköldkörtel. En blindsäck (cecum) som utgår från tarmen bakom gältarmen, anses vara en föregångare till ryggradsdjurens lever. Den försörjs nämligen, precis som levern, av en så kallad portåder, genom vilken den tillförs blod direkt från tarmens kapillärsystem.
Notera på den övre bilden att gälbågar och gälspringor är snedställda. Det är därför som de syns i tvärsnitt på den undre bilden. Färgade ljusmikroskopiska snitt. Modified images. Courtesy of John Houseman, from Wikimedia Commons under this GNU License. |
|
Gältarmen, gälbågarna och käkarna
Ett tredje kännetecken för ryggsträngsdjuren är gältarmen. Gältarmen är en svalgregion försedd med tvärgående springor, gälspringor, som avgränsas av gälbågar. Gältarmens ursprungliga funktion var sannolikt att filtrera födopartiklar från vattnet. Denna funktion har den fortfarande hos manteldjur och lansettfiskar, läs mer i bildtexten ovan. Eftersom den genomspolas av vatten, kan den samtidigt fungera som andningsorgan.
Manteldjur och lansettfiskar har ett mycket stort antal gälbågar. De första ryggradsdjuren var fiskar. Hos dem reducerades antalet gälbågar. De återstående gälbågarna försågs med skelett av brosk eller ben. Samtidigt försågs gältarmen med muskulatur och gälbågarna med ytförstorade utskott, gälar. Gältarmen fungerade därmed som ett andningsorgan med en muskeldriven pump, inte som en filtreringsapparat. Detta utvecklingsstadium finner man i princip hos de nutida käklösa ryggradsdjuren, pirålar och nejonögon, även om deras gälapparat är starkt modifierad.
Hos de käkförsedda ryggradsdjuren omvandlades först den främsta gälbågen, mandibularbågen, eventuellt flera främre gälbågar, till käkar med käkleder. Nästa gälbåge, hyoidbågen, utvecklades sedan till ett stöd för käkarna. De bakre fem gälbågarna bar gälar och ingick i andningsapparaten. Detta utvecklingsstadium finner man i princip hos alla nutida fiskar, utom pirålar och nejonögon. Uppkomsten av käkar var en revolution som gjorde det möjligt att utnyttja många nya födokällor.
Hos de fyrfota ryggradsdjuren modifierades käkarna, allra mest hos däggdjuren. Hos däggdjuren har den gamla käkleden, fantastiskt nog, ryckt in i mellanörat och en ny käkled utbildats. Läs om däggdjurens hörselben på en annan sida. Däggdjurens tungben (hyoidben) härstammar till största delen från hyoidbågen. Det är det enda större ben som saknar förbindelse med det övriga skelettet. Hos fyrfotadjuren uppträder gälbågarna under embryots utveckling och ger upphov till flera strukturer i halsregionen, men de tillbakabildas sedan. Läs mer gältarmens utveckling hos olika ryggradsdjur på en annan sida.
Hypostylen och sköldkörteln
Ett fjärde kännetecken för ryggsträngsdjuren är att de insöndrar jodhaltiga hormoner från särskilda celler. Hos lansettfiskar och manteldjur insöndras dessa hormoner från celler i hypostylen i gältarmens botten, se bilden ovan. Hos ryggradsdjuren bildar dessa celler en särskild körtel, sköldkörteln, som hos människan är belägen på halsen framsida. På en annan sida finns en bild av sköldkörteln. Där syns också bisköldkörtlarna. De härstammar från två par gälbågar. Det är därför som de är fyra stycken, ett par från varje gälbåge. Texten fortsätter under bilden.
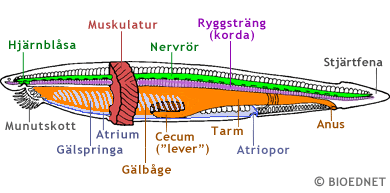 |
En schematisk översiktsbild av en lansettfisks anatomi. Större delen av muskulaturen har avlägsnat så att man kan se de inre organen. Det flesta av strukturerna är beskrivna i texten ovan. Notera dock ytterligare ett kännetecken för alla ryggsträngsdjur, nämligen att tarmen inte mynnar i bakänden. Anus är belägen framför den del av kroppen som kallas stjärt eller svans. Notera också atrioporen på undersidan. Det vatten som tas in genom munnen och passerar ut genom gälspringorna hamnar först i ett utanför tarmen beläget hålrum, atrium, och lämnar sedan kroppen via atrioporen. Modified image. Courtesy and copyright of BIODIDAC. |
|
Neurallist och plakoder
Två embryonala nybildningar har haft oerhört stor betydelse för ryggradsdjurens utveckling: neurallisten och plakoderna. Precis som nervröret, härstammar dessa strukturer från ektodermet, embryots yttersta cellskikt. Neurallisten utgörs av embryonala celler som uppkommer i det område, där nervröret snörps av. Dessa celler vandrar ut i hela embryot och utvecklas till en rad viktiga vävnader och celler. Läs mer om neurallisten på en annan sida. Lansettfiskarna bildar ingen neurallist. Manteldjuren gör det, men deras list ger bara upphov till pigmentceller. Plakoderna är förtjockningar av ektodermet. De ge bland annat upphov till luktorganen, ögonens linser samt inneröronens balansorgan inklusive hörselorganen. Lansettfiskar och manteldjur saknar plakoder. Haikouella och andra fossil från kambriumperioden har tolkats som föregångare till ryggradsdjuren eller tidiga ryggradsdjur med neurallist, men tolkningarna är osäkra.
Referenser
Texten har uppdaterats och utökats år 2014.
R. C. Brusca and G. J. Brusca: Invertebrates (Sinauer, 2nd ed, 2003).
J.-Y. Chen: Early crest animals and the insight they provide into the evolutionary origin of craniates (Genesis 46:623–639, 2008).
S. Conway Morris and J.-B. Caron: Pikaia gracilens Walcott, a stem-group
chordate from the middle Cambrian of British Columbia (Biological Reviews 87:480–512, 2012).
F. Delsuc, G. Tsagkogeorga, N. Lartillot, and H. Philippe: Additional molecular support for the new
chordate phylogeny (Genesis 46:592–604, 2008).
D. S. Fudge, N. Levy, S. Chiu, and J. M. Gosline: Composition, morphology and mechanics of hagfish slime (Journal of Experimental Biology 208:4613-4625, 2005).
J. Garcia-Fernàndez and È. Benito-Gutiérrez: It's a long way from amphioxus: descendants of the earliest chordate (BioEssays 31:665–675, 2009).
L. Z. Holland et al.: The amphioxus genome illuminates vertebrate origins and cephalochordate biology (Genome Research 18:1100-1111, 2008).
K.V. Kardong: The vertebrates (5th ed, McGraw-Hill, 2009).
Nicholas H. Putnam et al.: The amphioxus genome and the evolution of the chordate karyotype (Nature 453:1064-1072, 2008).
N. Takatori et al.: Comprehensive survey and classification of homeobox
genes in the genome of amphioxus, Branchiostoma floridae (Development, Genes and Evolution 218:579–590, 2008).
Till början på sidan
Till "Djurfakta"
|


