 |
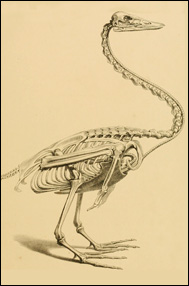 |
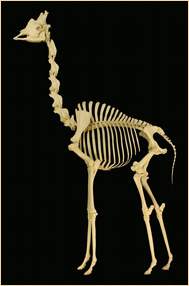
Giraffen (till vänster) har bara sju halskotor i den längsta halsen bland nu levande djur. Svanar (till höger) har 25 halskotor, fler än strutsen som har den längsta halsen bland nutida fåglar. Courtesy of the Museum of Osteology, Oklahoma City (left) and from T. C. Eyton et al." A monograph on the anatidae, or duck tribe" (1838), in the public domain. |
|
Nästan alla däggdjur har märkligt nog sju halskotor. Giraffens hals är förlängd, inte genom fler kotor, utan genom att varje kota är högre. Valarna har förlorat halsen som en anpassning till vattenliv, men har fortfarande sju förkortade halskotor, dock ofta sammanvuxna. Fåglar har dock ett mycket varierande antal halskotor.
Däggdjurens halskotor

De enda däggdjur som inte har sju halskotor är manaterna och sengångarna. Manaterna (släktet Trichechus) tillhör sjökorna, en grupp stora vattenlevande däggdjur, som varken är valar eller sälar. De har vanligen sex halskotor. Sengångarna lever i de tropiska delarna av Amerika och hänger upp och ner i träden med hjälp av sina förlängda klor. Enligt en vederhäftig källa uppges de tretåiga sengångarna (släktet Bradypus) har 8-10 halskotor, de tvåtåiga (släktet Choloepus) 5-6 hos en art och 7 hos en annan. Man hittar dock andra uppgifter om sengångarnas halskotsantal i litteraturen. Klart är emellertid att båda släktena kan ha ett halskotsantal som avviker från 7. De tretåiga sengångarna uppges kunna vrida sitt huvud upp emot tre kvarts varv (cirka 270 grader). Denna exceptionella rörlighet möjliggörs sannolikt av de övertaliga halskotorna. De tvåtåiga sengångarna uppges inte kunna vrida sitt huvud lika mycket. Texten fortsätter under bilden.
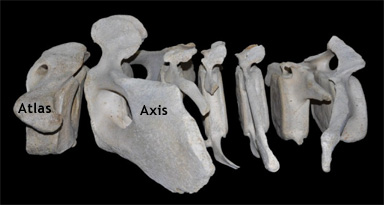 |
De sju halskotorna hos en sillval (Balaenoptera physalus). De två främsta halskotorna, axis och atlas, diskuteras längre ner på denna sida. Courtesy of E. Garcia Franquesa et al. (2014) under this Creative Commons License. |
|
Varför sju halskotor?
Varför har nästan alla däggdjur sju halskotor? Det är långt ifrån klarlagt, men det finns intressanta förslag. Halskotorna skiljer sig från bröstkotorna nedanför dem genom att de inte är anslutna till revben. Revben vid den nedersta halskotan förekommer som en ovanlig missbildning hos flera däggdjur, även hos människan. Detta innebär att denna kota förvandlats till en bröstkota. Missbildningen är kopplad till flera olika problem. Till dem hör en ökad risk att drabbas av cancer under de tidiga barnaåren. Det extra revbenet kan även utöva tryck på de nerver och den artär som försörjer armen. Detta kan ibland leda till att armen förtvinar. Möss med ett onormalt antal halskotor uppvisar en ökad cancerrisk. Allt detta borde leda till att individer med ett onormalt antal halskotor är mindre livsdugliga. Då borde också de genmutationer, som orsakar missbildningen, sorteras bort under evolutionen genom naturligt urval. En spekulativ förklaring till att manaterna och sengångarna kunnat utveckla ett avvikande halskotsantal är att dessa djur, på grund av sin låga energiomsättning, löper liten risk att drabbas av cancer.
Orsaken till halskoteabnormiteterna med de åtföljande negativa effekterna tros vara att en eller flera av däggdjurens Hox-gener inte fungerar normalt, något som stöds av molekylärbiologiska studier. Hox-generna styr embryots utveckling längs med kroppens längdaxel och därmed också anläggningen och utvecklingen av kotor och revben. Läs om Hox-genernas betydelse för ryggradsdjurens evolution på en annan sida.
Orsakerna till att manaterna och sengångarna har ett avvikande antal halskotor är inte klarlagda. De övertaliga halskotorna hos de tretåiga sengångarna skulle kunna tolkas så att gränsen mellan halsryggraden och bröstryggraden förskjutits bakåt under fosterutvecklingen. En alternativ möjlighet är att denna gräns inte förändrats. I stället har anlagen för vävnaderna vid sidan av ryggraden, det vill säga muskler och revben, förskjutits. Desssa anlag har i så fall flyttats bakåt hos tretåiga sengångare, så att 1-3 bröstkotor förlorat revben, och framåt hos tvåtåiga sengångare, så att 1-2 halskotor har fått revben. Även andra hypoteser har föreslagits. Texten fortsätter under bilden.
 |
Skelett av en tretåig sengångare (släktet Bradypus) med nio halskotor. Den är ett av mycket få däggdjur som inte har sju halskotor. Courtesy of Mike Taylor and the Cambridge University Museum of Zoology. |
|
Fåglarnas halskotor
Fåglar har ett mycket varierande antal halskotor. De restriktioner som finns hos däggdjur finns uppenbarligen inte hos dem. Det största antalet, 25 stycken, anges för svanar. När det gäller det minsta antalet anger en källa nio hos en del små fåglar, medan en annan anger elva hos en del papegojor. Förvånande nog nämndes strutsarna ingenstans, så jag gick upp på Zoologiska museet och kollade. Men strutsen (som finns i Afrika), emun och kasuarerna (stora strutslika fåglar i Australien) samt nanduerna (stora strutslika fåglar i Sydamerika) hade alla betydligt färre halskotor än svanarna.
De översta halskotorna: atlas och axis
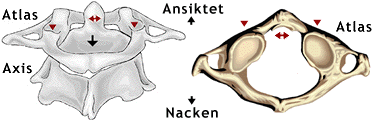
Hos både däggdjur och fåglar är den översta halskotan (atlas) och den näst översta (axis) speciellt utformade. Se bilden nedan. Därmed blir huvudet mera rörligt. När vi rör huvudet framåt och bakåt (d.v.s. nickar) använder vi lederna mellan kraniet och atlas. När vi snurrar på huvudet, som då vi säger nej, använder vi lederna mellan atlas och axis. Axis har en tapp som sticker upp inuti atlas. Rotationen sker kring denna tapp. När vi vaggar på huvudet i sidled, använder vi både lederna mellan kraniet och atlas och de mellan atlas och axis. Dessutom sker böjningar mellan de andra halskotorna när vi rör på huvudet. Hos valar, som ju inte rör på huvudet, kan atlas och axis eller till och med alla halskotorna vara sammanväxta. Texten fortsätter under bilden.
|
På den vänstra bilden syns människans översta nackkota, atlas, och näst översta, axis. Man ser kotorna snett uppifrån och bakifrån. På den högra bilden ser man atlaskotan uppifrån. Atlas och axis är utformade så att de ökar huvudets rörlighet och ser därför helt annorlunda ut än de andra ryggkotorna. De enkla röda pilspetsarna pekar på de två nedsänkta ledytor i atlas som ledar mot nackbenet i kraniet. De röda dubbelpilarna visar till vänster den tapp från axis som sticker upp inuti atlas och till höger den ledyta på atlas mot vilken tappen roterar när vi skakar på huvudet. Den svarta pilen på den vänstra bilden utpekar den kanal, genom vilken ryggmärgen löper i ryggraden. Se vidare texten nedan. Copyright 1996 © Corel Corporation. |
|
När vi nickar rör sig huvudet runt en rotationsaxel som går ungefär mellan de båda hörselgångarnas mynningar. Hos vuxna ligger huvudets tyngdpunkt framför denna axel. Det är därför som huvudet faller framåt, när man sitter i en stol och somnar. Hos
spädbarn är ansiktet mindre utvecklat, så huvudets tyngdpunkt ligger bakom rotationsaxeln. Det är därför som huvudet lätt faller bakåt hos spädbarn och måste stödjas när man håller barnet.
Atlaskotan har fått sitt namn efter den jätte i den grekiska mytologin som bar hela världen på sina axlar och därvid förstås måste luta huvudet framåt. Samme jätte har gett namn åt våra kartböcker.
Referenser
E. A. Buchholtz and C. C. Stepien: Anatomical transformation in mammals, developmental origin of aberrant cervical anatomy in tree sloths (Evolution & Development 11:69-79, 2009).
E. A. Buchholtz, K. L. Wayrynen, and I. W. Lin: Breaking constraint: axial patterning in Trichechus (Mammalia: Sirenia) (Evolution & Development 16:382-393, 2014).
F. Galis: Why do almost all mammals have seven cervical vertebrae? Developmental constraints, Hox genes, and cancer (Journal of Experimental Zoology [Mol Dev Evol] 285:19-26, 1999).
F. Galis and J. A. J. Metz: Testing the vulnerability of the phylotypic stage:
on modularity and evolutionary conservation (Journal of Experimental Zoology [Mol Dev Evol] 291:195-204, 2001).
E. Garcia Franquesa et al.: Descripción osteológica del rorcual común (Balaenoptera physalus, Linnaeus, 1758) del Museo de Ciencias Naturales de Barcelona (Arxius de Miscellània Zoològica Vol. 12, 2014).
L. Hautier et al.: Skeletal development in sloths and the evolution of
mammalian vertebral patterning (Proceedings of the National Academy of Science USA 107:18903-18908, 2010).
M.Taylor: Long and short-necked sloths of the Cambridge University Museum of Zoology (retrieved 20 October 2017).
I. Varela-Lasheras et al.: Breaking evolutionary and pleiotropic constraints in mammals, on sloths, manatees and homeotic mutations (EvoDevo 2:11, 2011).
Till början på sidan
Till "Djurfakta"
|


