 |
Skelettavgjutning av dinosaurien Apatosaurus (Brontosaurus) ett av världens största kända landdjur. Den levde under juraperioden (cirka 200-150 miljoner år före nutid). Notera att halsen hålls vågrät. De enormt tjocka skelettbenen i gångbenen tyder på att djuret kunde gå på land. Till synes har bara tåspetsarna markkontakt, åtminstone på frambenen. Men under tårna fanns troligen en stark elastisk bindvävsdyna, precis som hos dagens tyngsta landdäggdjur. Läs om skelettbenens tjocklek hos små och stora djur och om elefanternas klumpfötter på andra sidor. Läs mer om Apatosaurus anpassningar till sin kroppsvikt i faktarutan längst ner på denna sida. Courtesy of the University of Wyoming Museum and MCDinosaurhunter from Wikimedia Commons under this CC License. |
|
Förr trodde man att dinosaurier var tröga och långsamma djur med låg energiomsättning, i motsats till dem, som man trodde, "högre" utvecklade däggdjuren. Men man hade fel. Dinosaurierna dominerade trots allt bland de större landlevande djuren under betydligt längre tid än vi däggdjur har gjort det. Och de är inte utdöda. Fåglarna är nu levande dinosaurier. Dessutom var de utdöda dinosaurierna sannnolikt inte "kallblodiga", alltså växelvarma.
Växelvarma och jämnvarma djur

Man trodde således förr att de utdöda dinosaurierna, precis som de flesta av dagens reptiler, var poikiloterma (växelvarma, "kallblodiga"), d.v.s. höll samma temperatur som omgivningen, och att de var ektoterma, d.v.s. inte kunde värma upp sig med egenproducerad kroppsvärme. Man trodde också att de inte kunde reglera sin kroppstemperatur, förutom genom beteende, till exempel genom att värma upp sig med hjälp av solvärme.
Nästan alla däggdjur och fåglar är homeoterma (jämnvarma, "varmblodiga"), d.v.s. håller en nära nog konstant kroppstemperatur, och endoterma, d.v.s. kan värma upp sig med egenproducerad kroppsvärme. De kan också reglera sin kroppstemperatur på en nära nog konstant nivå via andra mekanismer än beteende.
Var dinosaurierna jämnvarma?
Numera har man drastiskt ändrat sin uppfattning om de utdöda dinosaurierna. De flesta menar att de kunde värma upp sig själva och troligen också reglera sin kroppstemperatur. Men hade de en lika hög inre värmeproduktion och en lika effektiv temperaturreglering som däggdjur och fåglar? Här går åsikterna isär. Det handlar ju om fysiologiska egenskaper och sådana fossileras inte. Dessutom har man mycket få fossil med bevarade mjukdelar. Vanligen har man bara skelettdelar. Vilka bevis tyder då på att de utdöda dinosaurierna var homeoterma, endoterma reglerare?
Man vet nu att fåglarna är en gren på dinosauriernas släktträd. Man kan därför misstänka att de andra dinosaurierna hade många egenskaper gemensamma med fåglarna. Men detta är inte hårda fakta. Texten fortsätter under bilderna.
 |
 |
Fotspår av två dinosaurier, till vänster av en tyrannosauriesläkting och till höger av en apatosaurussläkting, en sauropod. Spåret av den lättare tyrannosauriden påminner mycket om ett fotspår av en fågel, till exempel en struts, med tårna nummer II-IV framåtriktade. Spåret av den betydligt tyngre sauropoden ser ut precis som ett spår av en elefants klumpfot. Foten är alltså anpassad till att bära upp djurets enorma tyngd. Courtesy of R. T. McCrea et al. from "A ‘Terror of Tyrannosaurs’: the first trackways of tyrannosaurids and evidence of gregariousness and pathology in Tyrannosauridae" (PLoS ONE 9[7]: e103613, 2014; left) and T. Thulborn from "Impact of sauropod dinosaurs on lagoonal substrates in the Broome sandstone (lower Cretaceous), Western Australia"(PLoS ONE 7[5]: e36208, 2012; right), both under this CC License. |
|
Hög tillväxthastighet
De fossila dinosaurieägg man hittat är förvånande små i förhållande till djurens storlek. Däremot var de nykläckta ungarnas tillväxthastighet förvånande hög. Man har kunnat mäta tillväxthastigheten genom att räkna årsringar i skelettbenen och uppskatta djurens kroppsvikt utifrån benens storlek. Alla undersökta utdöda dinosaurier av olika storlek växte betydligt snabbare än nutida kräldjur. Stora dinosaurier hade tillväxthastigheter i samma storleksordning som nutida däggdjur och fåglar. Denna snabba tillväxt tyder på att de växande dinosaurierna hade en hög energiomsättning och därmed en hög inre värmeproduktion, alltså endotermi.
Stora djur kan lättare hålla sig varma
De stora dinosaurierna var så stora att de sannolikt kunde höja sin kroppstemperatur över omgivningens temperatur, även med en förhållandevis liten värmeproduktion. Detta beror på att större djur har en mindre värmeavgivande kroppsyta mätt per kilo kroppsvikt. Fenomenet kallas gigantotermi. Man har påvisat gigantotermi hos några nu levande stora reptiler, nämligen hos stora krokodiler och hos havslädersköldpaddan. Havslädersköldpaddan kan också till en viss del reglera sin kroppstemperatur, åtminstone när den simmar. Läs mer om kroppsviktens och kroppsytans betydelse för däggdjurs energiomsättning och värmeproduktion på en annan sida.
Riskerade stora dinosaurier överhettning?
Till detta kommer att klimatet under dinosauriernas tidsålder var varmare än nu. Det var kanske till och med så att mycket stora dinosaurier hade blivit överhettade, om de hade samma höga energiomsättning som dagens däggdjur och fåglar. Nutida elefanter måste ha en rad anpassningar som motverkar överhettning. Och de är betydligt mindre än de största dinosaurierna. Kanske var det så att dinosauriernas snabbväxande ungar hade en hög energiomsättning, men att mycket stora vuxna dinosaurier måste ha haft en betydligt lägre omsättning än däggdjur och fåglar.
Tecken på hög kroppstemperatur
Uppskattningar av dinosauriernas kroppstemperaturer tyder på att de var endoterma, men ger olika resultat. I en studie mätte man isotoper av kol och syre i fossila tänder från stora dinosaurier. Denna studie tyder på att dessa djur hade en kroppstemperatur på 36-38 °C, ungefär som dagens däggdjur. I en annan studie, baserad på tillväxthastighet, kom man fram till en kroppstemperatur på 25 °C för en dinosaurie på 12 kg och 41 °C för en dinosaurie på 13 ton. Man har också gjort fossilfynd, där många individer av flera olika dinosauriearter har hittats tillsammans. I dessa fynd tycks andelen köttätande relativt växtätande individer vara närmare den som gäller för nutida däggdjur än den som gäller för nutida kräldjur. Detta tyder på att de utdöda dinosaurierna hade en hög energiomsättning och därmed en hög inre värmeproduktion, med andra ord att de var endoterma. Dessutom fanns det vissa dinosaurier i polartrakterna, där inga av dagens reptiler kan överleva. Dessa dinosaurier hade knappast kunnat överleva där utan att hålla kroppens temperatur högre än omgivningens. De hade förmodligen också en isolerande fjäderdräkt. Texten fortsätter under bilden.
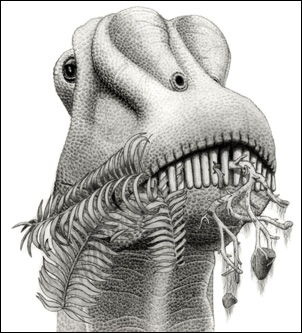 |
Ett konstnärligt porträtt av dinosaurie, en släkting till Diplodocus. Notera att den käkar grenar av barrträd. Då den levde, hade blomväxterna ännu inte erövrat världen. Födan var svårspjälkad. Precis som många fåglar gör i dag, svalde många växtätande utdöda dinosaurier stenar, så kallade gastroliter, som i muskelmagen hjälpte till att mekaniskt byta ner födan. Courtesy of Jordan Mallon under this CC License. |
|
Dinosauriernas hjärta och lungor
Man har länge frågat sig om de utdöda dinosaurierna hade ett trerummigt hjärta, som dagens kräldjur, eller ett fyrrummigt, som dagens däggdjur och fåglar. Problemet troddes vara löst, när man hittade ett bevarat fossilt hjärta. Detta beskrevs som fyrrummigt med en enda aortabåge, precis som däggdjurens och fåglarnas. Fyndet har dock sedermera starkt ifrågasatts. Icke desto mindre tror dock de flesta att de utdöda dinosaurierna hade ett fyrrummigt fågelliknande hjärta. Ett skäl är att långhalsade dinosaurier inte hade kunnat producera ett tillräckligt högt blodtryck för att förse hjärnan med syrerikt blod, om de hade haft ett trerummigt hjärta. För en utförligare förklaring, läs om trerummiga och fyrrummiga hjärtan på en annan sida.
Många skelettben hos utdöda dinosaurier hade stora inre hålrum och var därmed förvånansvärt lätta. Hålrummens struktur tyder på att de var fyllda med luft, precis som hos fåglar. Hos fåglarna är de luftfyllda hålrummen delar av de så kallade luftsäckarna. Luftsäckarna ingår i fåglarnas andningsapparat, som ger dem ett avsevärt effektivare syreupptag än däggdjurens. Revbenens struktur hos flera utdöda dinosaurier tyder på att de hade en fågelliknande andningsmekanism. Allt detta stödjer tanken att de utdöda dinosaurierna hade en liknande andningsapparat som fåglarna. De kan därför ha haft ett stort syreupptag, vilket möjliggjorde en hög energiomsättning och värmeproduktion, alltså endotermi. Läs om hur fåglar andas på en annan sida. Texten fortsätter under faktarutan.
|
Om hur lungornas funktion och globala katastrofer kan ha påverkat dinosauriernas och däggdjurens evolution
Under karbonperioden och början av permperioden var syrehalten i atmosfären upp emot 30 %, mycket högre än dagens 21 %. Med början under perm för cirka 300 miljoner år sedan sjönk sedan halten för att under triasperioden (ca 250-200 miljoner år före nutid) nå en bottennivå på cirka 10 %. Vid övergången mellan perm och trias ledde dessutom globala katastrofer till det största massutdöendet av djurarter någonsin. De dominerande större landdjuren under perm var så kallade synapsida reptiler, bland dem föregångare till dagens däggdjur. De flesta av dem dog ut.
Under trias skedde i stället en snabb utveckling av dinosaurier. De fyllde de lediga ekologiska nischerna och blev nu de dominerande större landdjuren. Man tror att detta åtminstone delvis berodde på att dinosaurierna hade en fågelliknande andningsapparat och därmed lättare klarade av luftens låga syrehalt. En del synapsider klarade sig dock och gav så småningom upphov till däggdjuren. Men detta skedde först efter det stora utdöendet vid meteoritnedslaget för cirka 70 miljoner år sedan. Då ökade däggdjuren kraftigt i antal. Eftersom alla dinosaurier utom fåglarna hade dött ut, kunde däggdjuren ta över en mängd tomma nischer. Syrehalten i luften låg då på ungefär samma nivå som nu. |
|
Inte växelvarma, inte jämnvarma, men mitt emellan?
Hos nu levande djur finns det ett matematiskt samband mellan tillväxthastighet och energiomsättning i vila. I en studie från 2014 använde man detta samband till att uppskatta energiomsättningen i vila hos flera utdöda dinosaurier. Man uppskattade också djurens kroppsvikt utifrån skelettbenens dimensioner. Sedan konstruerade man ett diagram som jämförde energiomsättning och kroppsvikt hos de utdöda dinosaurierna med ett stort antal nutida djur, både jämnvarma och växelvarma.
De utdöda dinosaurierna hamnade i diagrammet mellan de jämnvarma och de växelvarma djuren. Man drog slutsatsen att dinosaurierna hade en medelhög energiomsättning och att deras kroppstemperatur var högre än omgivningens temperatur, men inte lika hög som hos nutida jämnvarma däggdjur och fåglar. Man antog att de hade en varierande kroppstemperatur, som reglerades inom vida gränser eller inte reglerades alls. Detta mellanläge mellan endotermi och ektotermi kallade man mesotermi. Vissa nutida djur klassificerades också som mesoterma, bland annat vissa fiskar, den ovan nämnda havslädersköldpaddan samt, bland däggdjuren, kloakdjuren. Väger man samman den information som finns, förefaller detta vara en tilltalande hypotes. Å andra sidan borde det rimligen ha funnits stora skillnader i temperaturanpassning mellan olika grupper av dinosaurier. Läs om mesoterma fiskar och om mesoterma däggdjur på andra sidor.
Var fisködlor och flygödlor också jämnvarma?
Det finns data som tyder på att inte bara dinosaurierna, utan också de utdöda fisködlorna (ichthyosaurierna) och flygödlorna (pterosaurierna), var jämnvarma. Dessa grupper var inga ödlor och inte heller dinosaurier. De var kräldjur evolutionärt anpassade till simning respektive flygning och var inte nära släkt med andra reptilgrupper. De var samtida med dinosaurierna.
Fisködlorna är ett klassiskt exempel på så kallad konvergent evolution, det vill säga oberoende utveckling av liknande anpassningar hos obesläktade djur med likartat levnadssätt. De påminde mycket om delfiner och var liksom de senare marina snabbsimmande fiskätare. Till likheterna hörde kroppsformen, gångben omvandlade till fenor samt tanduppsättningen. Fisködlorna födde också, precis som delfinerna, levande ungar. Till skillnad från delfinerna hade fisködlorna dock bakben omvandlade till fenor samt en lodrät, inte en vågrät, stjärtfena. Texten fortsätter under bilderna.
 |
 |
Överst ses ett fossil av en fisködla. Notera likheten med en delfin. Notera också att mjukdelar är bevarade. Man se var huden avgränsar kroppen. Stjärtfenans övre flik och ryggfenan saknar helt skelett. Ryggraden gör en tvär böj ner i stjärtfenans undre flik. Nederst ses en teckning av en fisködlehona som föder en unge. Den är baserad på fossilfynd. Ungen föds med stjärten först, precis som hos valar. Detta möjligör troligen syretillförsel via en navelsträng under själva födseln, innan ungen kan nå vattenytan och andas med lungorna. Så är det hos valar. Normalt föder däggdjur sina ungar med huvudet först. Inuti modern ovan finns ytterligare tre ungar, som väntar på att födas. Valar föder alltid bara en unge.
I ögats senhinna hade fisködlorna en stödjande ring av små ben, som syns på bilden. Vissa fisködlor hade jättestora ögon, kanske de största man känner till. Större ögon släpper in mer ljus och ger därmed ett bättre mörkerseende. Detta tyder på att dessa fisködlor var goda dykare och kunde dyka mycket djupt ner i havet, dit mycket lite ljus når. Courtesy of Staatliches Museum für Naturkunde (Stuttgart) in the public domain (above) and Motani et al. under this CC License (below). |
|
I en studie från 2018 påvisade man ytterligare likheter med delfinerna hos ett särskilt välbevarat fisködlefossil. Dess hudyta var jämn utan fjäll och innehöll ett späcklager, precis som hos delfiner. Späck gör kroppen mer strömlinjeformad och ökar dess flytförmåga. Särskilt intressant är att späcket hos dagens valar och sälar ger värmeisolering. Därmed förhindras allt för stora värmeförluster till det omgivande vattnet, som har mycket hög värmeledningsförmåga. En låg värmeförlust är nödvändig för de vattenlevande djur som är jämnvarma ("varmblodiga") och håller en nära nog konstant kropptemperatur. Späcket tyder på att fisködlorna kanske var jämnvarma, precis som däggdjur och fåglar.
Fördelningen av pigmentkorn i fisködlefossilets hud tyder på att fisködlor, precis som delfiner, var mörka på översidan och ljusa på undersidan. Detta ger kamouflage, eftersom djuren ser vita ut underifrån mot den ljusa vattenytans bakgrund och svarta ovanifrån mot det mörka havsdjupets bakgrund. Fisködlans pigmentceller liknade däremot inte delfinernas, utan var stjärnformade som hos många reptiler. Möjligen kunde djuren skifta färg, precis som många ödlor. Läs om färgväxling och om späck på andra sidor.
På flygödlornas hud har man i väl bevarade fossil identifierat grenade eller ogrenade fibrer, som skulle kunna vara en slags pälsliknande kroppsbeklädnad. Dessa fibrer har tolkats som primitiva fjädrar, men detta är kontroversiellt. I vilket fall som helst, skulle fibrerna kunna fungera som värmeisolering, precis som fåglarnas fjädrar och däggdjurens hår. Dett tyder på att flygödlorna möjligen var jämnvarma djur. Läs mer om flygödlor på en annan sida.
|
 |
 |
Sauropoderna, världens största kända landdjur någonsin
Till dinosauriegruppen Sauropoda hör de tyngsta kända landdjuren någonsin. Uppfattningen om deras levnadssätt har drastiskt förändrats genom åren. Överst ses en konstnärlig rekonstruktion från början av 1900-talet av sauropoden Apatosaurus (Brontosaurus). Det ljusare djuret, som står på land är dock en Diplodocus. Uppskattningen av Apatosaurus vikt är osäkra, men värden från cirka 20 till cirka 50 ton uppges ofta. Man trodde förr att Apatosaurus var ett "kallblodigt" trögt djur, som var så tungt att det inte kunde bära sin egen kropp på land utan måste leva i vatten.
Notera emellertid djuren i bakgrunden som bara sticker upp huvudet över vattnet. Ett rimligt antagande är då att avståndet mellan vattenytan och lungorna är cirka 10 meter. På detta djup är vattnets tryck cirka 2 atm, dubbelt så högt som i luften ovanför. Trycket inuti bröstkorgens vävnader, inklusive lungans blodkapillärer, är då cirka 1 atm högre än på land. Eftersom lungorna är förbundna med luften via luftvägarna, är lufttrycket inuti lungorna lika med lufttrycket i atmosfären ovanför vattenytan, cirka 1 atm. Med en tryckskillnad på cirka 1 atm mellan vävnaderna och lungluften skulle lungkapillärerna brista och lungan fyllas med blod. Dessutom skulle djuret inte kunna utvidga lungorna för att andas in. Vattensamlingen skulle fyllas av döda apatosaurier.
Nederst ses en modern rekonstruktion av en annan sauropod, Brontomerus. Numera anser man att sauropoderna var mer eller mindre "varmblodiga" (jämnvarma) och att de kunde röra sig på land. De skulle till och med kunna ha varit så alerta, att honan på bilden kunde sparka ihjäl den rovdinosaurie, som hotar hennes unge. Man anser numera också att sauropoderna normalt höll halsen vågrät eller endast något upphöjd i förhållande till kroppen. Utformningen av vissa sauropoders halskotor tyder på att de inte kunde höja halsen. Om de kunde höja halsen till lodrät position, kunde de förmodligen inte göra detta under någon längre tid. Då skulle hjärnans blodkärl kollapsa på grund av undertryck i dem relativt atmosfärstrycket. Alternativt skulle halsen kunna ha varit försedd med ett antal hjälphjärtan, som höjde blodtrycket till en tillräckligt hög nivå. För att få en förklaring till tyngdkraftens effekter på blodomloppet, läs om hur giraffen klarar av att försörja hjärnan med blod på en annan sida. Images by Charles Robert Knight, in the public domain (above) and by Francisco Gascó under this Creative Commons License (below). |
|
Referenser
M. Balter: Dinosaur metabolism neither hot nor cold, but just right (Science 344:1216-1217, 2014).
B. L. Bostrom and D. R. Jones: Exercise warms adult leatherback turtles (Comparative Biochemistry and Physiology A 147:323-331, 2007).
J. P. Casey, M. C. James, and A. S. Williard: Behavioral and metabolic contributions to thermoregulation in freely swimming leatherback turtles at high latitudes (Journal of Experimental Biology 217:2331-2337, 2014).
R. A. Eagle et al.: Dinosaur body temperatures determined from isotopic (13C-18O) ordering in fossil biominerals (Science 333:443-445, 2011).
G. M. Erickson, K. C. Rogers, and S. A. Yerby: Dinosaurian growth patterns and rapid avian growth rates (Nature 412:429-433, 2001).
P. E. Fisher et al.: Cardiovascular evidence for an intermediate or higher metabolic rate in an ornithischian dinosaur (Science 288:503-505, 2000).
J. F. Gillooly , A. P. Allen, and E. L. Charno: Dinosaur fossils predict body temperatures (PLoS Biol 4[8]: e248, 2006).
John M. Grady et al.: Evidence for mesothermy in dinosaurs (Science 344:1268-1272, 2014).
J. Lindgren et al.: Soft-tissue evidence for homeothermy and crypsis in a Jurassic ichthyosaur (Nature 564:359-365, 2018).
R. Motani et al.: Terrestrial origin of viviparity in Mesozoic marine reptiles Indicated by early triassic embryonic fossils (PLoS ONE 9: e88640, 2014).
F. V. Paladino, M. P. O'Connor, and J. R. Spotila: Metabolism of leatherback turtles, gigantothermy, and thermoregulation of dinosaurs (Nature 344:858-860, 1990).
D. J. Pierson: The physiology of dinosaurs, circulatory and respiratory function in the largest animals ever to walk the earth (Respiratory Care 54:887-911, 2009).
G. D. Ruxton: Heat loss from giant extinct reptiles (Proceedings of the Royal Society London B 268:1921-1924, 2001).
P. M. Sander et al.: Adaptive radiation in sauropod dinosaurs: bone histology indicates
rapid evolution of giant body size through acceleration (Organisms, Diversity & Evolution 4:165-173, 2004).
P. C. Sereno et al.: Evidence for avian intrathoracic air sacs in a new predatory dinosaur from Argentina (PLoS ONE 3[9]: e3303, 2008).
M. P. Taylor et al.: Brontomerus mcintoshi, a new sauropod dinosaur from the lower Cretaceous Cedar Mountain formation, Utah, USA (Acta Palaeontologica Polonica 56:75-98, 2011).
Z. Yang et al.: Pterosaur integumentary structures with complex feather-like branching (Nature Ecology & Evolution 3:24-30, 2019).
Till början på sidan
Till "Djurfakta"
|


